
With the recent NIH requirement to include female subjects in biomedical research, you may be wondering how this will complicate your experiments. Adding a female cohort already doubles the required number of experimental animals, but hormonal fluctuations due to the female reproductive cycle will add an additional variable. Should reproductive cycles be considered in behavioral research, and why?
The female reproductive cycle, called the estrous cycle in rodents, involves cyclical changes in hormones such as estrogen, progesterone, and their metabolites. Estradiol, the main circulating estrogen, reaches a peak mid-cycle that triggers ovulation, after which progesterone is released in high levels from the ovary. If fertilization does not occur, hormone levels fall rapidly at the conclusion of the cycle
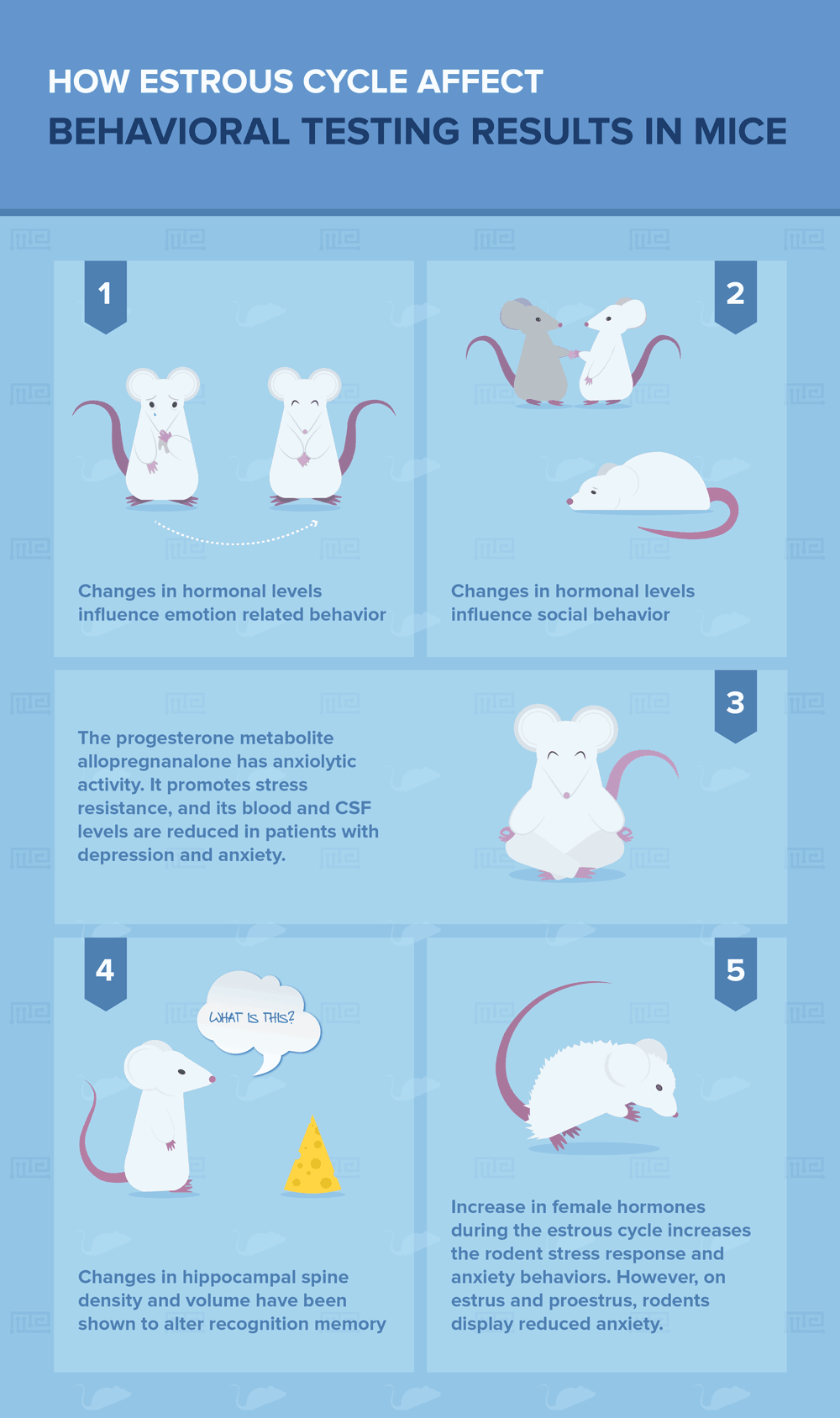
These changing levels of hormones have been demonstrated to influence a variety of animal behaviors, including emotion-related behaviors, social behaviors, drug taking, and cognition, among others[1]. These effects are largely caused by the direct actions of reproductive hormones in the brain, which expresses steroid receptors throughout behaviorally relevant brain regions.
Steroid derivatives can also affect brain activity. For instance, the progesterone metabolite allopregnanalone acts as an agonist at GABAA receptors, the major inhibitory receptors in the brain. Allopregnanolone has anxiolytic activity and promotes stress resistance. Its blood and CSF levels are reduced in patients with depression and anxiety[3, 4].
With respect to effects on cognition, estradiol fluctuations across the estrous cycle are associated with changes in hippocampal spine density and volume[2, 6]. These changes have been shown to alter recognition memory and are associated with the use of different navigation strategies in the T maze[5].
Females also show cyclical changes in tests of anxiety and depression. The increase in female hormones during the estrous cycle increases the rodent stress response and anxiety behaviors[6]. However, on estrus and proestrus (the ovulatory stage of the cycle), rodents display reduced anxiety[8, 9]. Depression-like behavior in the forced-swim test also shows estrous cycle-dependent changes[7].
These data clearly demonstrate the importance of considering this variable in your behavioral experiments. How can cycle effects be controlled?
Infographic Illustrates How Estrous Cycle Affect Behavioral Testing Results In Mice
There are several possibilities for either assessing or minimizing the impact of reproductive cycles on experimental outcomes. All of these methods require tracking the cycles. In rodents, estrous cycles are monitored using vaginal cytology, which involves taking vaginal swabs and using a microscope to analyze the vaginal epithelial cells, which show characteristic changes during each cycle stage.
To minimize the effects of hormones on behavior, females should be studied during the phase of the cycle in which hormonal changes are the least dynamic, which is diestrus in rodents. In behavioral tests where there isn’t a large overall sex difference, diestrus females behave most similarly to males. The difficulty of this method is that your experimental schedule will be determined by the animals’ cycles, which can vary in length and timing.
You can also choose to run your experiments as scheduled, assess the estrous cycle stage after testing each animal, and then run statistical tests to assess possible effects of cycle stage. However, without a large number of animals, you will have a limited statistical power to detect differences, since relatively few animals are likely to be in proestrus on any given day.
If you are interested in studying rather than eliminating a potential effect of cycles on behavior, you can track the animals’ cycles and then administer tests to a specific number of mice per cycle stage. However, this method will be the most time consuming, as it may take days to weeks to obtain the appropriate number of animals in proestrus.
References
- ter Horst JP, de Kloet ER, Schächinger H, Oitzl MS. (2012) Relevance of stress and female sex hormones for emotion and cognition. Cell Mol Neurobiol. 32(5):725-35.
- Marques AA, Bevilaqua MC, da Fonseca AM, Nardi AE, Thuret S, Dias GP. (2016) Gender Differences in the Neurobiology of Anxiety: Focus on Adult Hippocampal Neurogenesis. Neural Plast. 2016:5026713.
- Yoshizawa K, Okumura A, Nakashima K, Sato T, Higashi T. (2017) Role of allopregnanolone biosynthesis in acute stress-induced anxiety-like behaviors in mice. Synapse. 2017 Apr. (epub ahead of print)
- Schüle C, Nothdurfter C, Rupprecht R. (2014) The role of allopregnanolone in depression and anxiety. Prog Neurobiol. 113:79-87.
- Qiu LR, Germann J, Spring S, Alm C, Vousden DA, Palmert MR, Lerch JP. (2013) Hippocampal volumes differ across the mouse estrous cycle, can change within 24 hours, and associate with cognitive strategies. Neuroimage. 83:593-8.
- Luine V, Frankfurt M. (2013) Interactions between estradiol, BDNF and dendritic spines in promoting memory. Neuroscience. 239:34-45.
- Kokras N, Antoniou K, Mikail HG, Kafetzopoulos V, Papadopoulou-Daifoti Z, Dalla C. (2015) Forced swim test: What about females? Neuropharmacology. 99:408-21.
- Ramos-Ortolaza DL, Doreste-Mendez RJ, Alvarado-Torres JK, Torres-Reveron A. (2017) Ovarian hormones modify anxiety behavior and glucocorticoid receptors after chronic social isolation stress. Behav Brain Res. 328:115-122.
- Borrow AP, Cameron NM. (2017) Maternal care and affective behavior in female offspring: Implication of the neurosteroid/GABAergic system. Psychoneuroendocrinology. 76:29-37.