



In the world of behavioral neuroscience, precision is no longer a luxury—it’s a necessity. As research evolves from broad descriptions of behavior to fine-grained, quantifiable metrics, scientists are increasingly seeking tools that go beyond traditional endpoints like escape latency or quadrant dwell time. Nowhere is this shift more evident than in the Morris Water Maze, a cornerstone of spatial memory research that, despite its apparent simplicity, holds a vast reservoir of untapped behavioral insight.
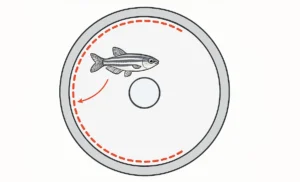
One such insight lies in a spatial construct known as Whishaw’s Corridor—an invisible path linking the animal’s start location to the platform. While invisible to the naked eye, this narrow corridor becomes a powerful behavioral lens when paired with modern tracking technology. It provides a refined framework to assess not just whether an animal finds the platform, but how accurately, how efficiently, and with what degree of cognitive certainty it navigates the space.
At the heart of this analysis is distance travelled within Whishaw’s Corridor—a metric that quantifies goal-directed behavior with remarkable sensitivity. It captures nuances of spatial memory, decision-making strategies, and confidence in recall that conventional measures may overlook. With this metric, a maze trial becomes more than a binary test of success or failure—it becomes a behavioral narrative, one that reveals the quality of memory and the structure of strategy.
As demands for reproducibility, translational relevance, and behavioral depth grow, so too must our approaches. Whishaw’s Corridor offers a bridge between spatial behavior and neural computation—anchored in hippocampal function and expressive of behavioral integrity. It’s a metric that aligns with the future of neuroscience: precise, contextual, and insight-rich.
In the realm of behavioral neuroscience, where the goal is to disentangle purposeful navigation from chance-driven motion, Whishaw’s Corridor stands out as a powerful spatial analysis tool. Conceptualized by Dr. Ian Q. Whishaw, this “corridor” is not a physical structure in the Morris Water Maze, but a virtual, experimenter-defined path—a rectangular channel extending from the subject’s start location directly to the goal platform.
Functionally, it represents the most efficient, ideal route that a rodent would take if it possessed perfect knowledge of the platform’s location. In essence, Whishaw’s Corridor is a behavioral hypothesis made spatial: it proposes a zone of goal-directed action and allows researchers to test how closely the subject’s actual swim behavior conforms to this optimal trajectory.
The corridor becomes especially meaningful during probe trials, where the platform is removed to test memory retention. If a rodent’s swim path remains within or near the confines of Whishaw’s Corridor, it suggests strong spatial encoding and the use of allocentric navigation strategies—those relying on distal environmental cues, which are primarily mediated by the hippocampus.
More than just a line of best fit, Whishaw’s Corridor serves as a metric of intentionality. By tracking the animal’s proximity to the corridor and measuring the distance travelled within it, researchers can evaluate not just whether the animal remembered the platform’s location, but how confidently and precisely it acted upon that memory.
This distinction is critical. A rodent may eventually find the platform through procedural tactics like thigmotaxis or serial scanning, but such behaviors reflect non-cognitive problem solving, rather than true spatial learning. In contrast, repeated and consistent navigation within Whishaw’s Corridor reflects cognitive mapping—the internal construction of space that allows flexible, efficient recall and decision-making.
With the integration of high-resolution video tracking and automated path analysis, this theoretical construct becomes quantifiable. Modern tools, like those provided by Conduct Science, can digitally map the corridor and overlay the subject’s swim trace, calculating distance, deviation, and efficiency in real time.
In this way, Whishaw’s Corridor transforms behavioral data from a basic observation into a precise neuroscientific measurement, illuminating the distinction between success through memory and success through repetition.
While traditional metrics in the Morris Water Maze—such as escape latency or quadrant dwell time—offer coarse indicators of task performance, they often obscure the finer details of how an animal reaches the platform and what that reveals about its cognitive processes. This is where distance travelled in Whishaw’s Corridor offers exceptional value: it captures a rodent’s spatial precision and the clarity of its internal navigational map.
Whishaw’s Corridor isn’t just a path—it’s a behavioral benchmark. Movement through this corridor reflects a high-fidelity execution of spatial memory. When a rodent swims within this virtual channel, it is not simply demonstrating that it knows where the platform is—it is behaving as though it is certain of its location. The more tightly a swim path adheres to the corridor, the more confidently we can infer that the animal is using allocentric spatial cues to guide its navigation.
This is especially important in distinguishing between strategic learning and chance performance. For example, a rodent may reach the platform quickly by relying on procedural habits (such as hugging the wall or scanning the pool) rather than true memory recall. These strategies can result in low escape latency, but high variability and unnecessary distance travelled—often outside the corridor. In contrast, when an animal’s swim path conforms tightly to Whishaw’s Corridor and the total distance travelled within it decreases over trials, it signals a refinement of cognitive strategy and increased navigational certainty.
This behavioral certainty is more than just task familiarity—it reflects a neurobiological commitment to the goal. It is the behavioral manifestation of hippocampal encoding, where place cells, grid cells, and path integration mechanisms converge to form an internal representation of the environment (O’Keefe & Nadel, 1978). When that representation is strong, the animal does not explore—it acts.
Furthermore, Whishaw’s Corridor provides a unique opportunity to assess the efficiency of learning over time. During early trials, even healthy control animals may deviate from the corridor as they form initial spatial associations. But as training progresses, successful learners show a consolidation effect: their swim paths become straighter, shorter, and more aligned with the corridor. This convergence reflects not only memory acquisition but behavioral optimization—a hallmark of cognitive mastery.
Equally important is how this measure performs under conditions of impairment. In rodent models of Alzheimer’s disease, stroke, or traumatic brain injury, animals may still reach the platform but exhibit significant deviation from the corridor and travel greater distances within it than their healthy counterparts. This increased distance is not due to weakness or motor impairment, but to impaired spatial processing and decision-making noise. The animal’s search may appear aimless or inefficient, signaling uncertainty in memory retrieval or attentional control.
By capturing these nuances, distance in Whishaw’s Corridor becomes a proxy for cognitive clarity—one that transcends binary task success and delves into the quality of the behavior itself. It tells us not only that the animal reached its goal, but how directly, confidently, and accurately it did so.
At Conduct Science, we recognize that these distinctions matter. Our behavioral analysis platforms are designed to help researchers move beyond surface-level measures, enabling them to trace behavioral intention, evaluate treatment effects, and dissect learning trajectories with granular resolution. Because in neuroscience, it’s not just about reaching the goal—it’s about understanding the path taken to get there.
The behavioral metric of distance travelled within Whishaw’s Corridor has evolved into a robust, sensitive measure for detecting cognitive and neurological dysfunction across a range of preclinical disease models. In neuroscience research, especially using the Morris Water Maze, subtle deficits in memory or attention often go unnoticed if relying solely on escape latency or platform crossings. This is where the corridor-based analysis becomes essential—it captures not just whether an animal reaches the goal, but how it gets there.
In models of Alzheimer’s disease, for example, rodents often exhibit increased corridor distance, not because they are physically unable to swim, but because they lack the spatial precision required to move directly to the platform. They may rely on procedural or egocentric strategies—like wall-following or random scanning—resulting in elongated, inefficient paths. This increase in distance within or deviating from the corridor becomes a behavioral fingerprint of hippocampal dysfunction.
Similarly, in traumatic brain injury (TBI) or stroke models, animals typically show impaired path straightness and longer intra-corridor distances even after initial motor recovery. These deviations suggest disrupted spatial processing and diminished cue integration, despite preserved gross mobility. As shown in the graph above, TBI and Alzheimer’s model groups tend to travel significantly farther within Whishaw’s Corridor compared to healthy controls.
Moreover, genetic knockout (KO) models—such as mice lacking NMDA receptor subunits or key neuroplasticity genes—also show performance degradation in this metric. Their paths may appear erratic or unfocused, with longer corridor distances that reflect both reduced learning consolidation and increased behavioral variability.
In contrast, pharmacological intervention groups—such as rodents treated with cholinergic enhancers, NMDA modulators, or neuroprotective agents—often exhibit partial or full restoration of spatial efficiency. This is seen as a reduction in average distance travelled within the corridor, moving the profile closer to that of control animals.
The bar chart provided illustrates how average corridor distance varies across experimental conditions. Control animals typically travel the shortest distances—demonstrating learned, efficient navigation. Disease models show elongated distances, while treatment groups show improvements that can be tracked quantitatively.
This metric serves as both a diagnostic indicator and a therapeutic readout, making it indispensable in preclinical trials aiming to reverse or mitigate cognitive decline. The ability to continuously monitor distance within Whishaw’s Corridor across training sessions also provides a powerful longitudinal measure, capturing the dynamics of learning and recovery over time.
At Conduct Science, our maze tracking systems and behavior analytics software are specifically designed to extract, visualize, and interpret these subtle but significant metrics—helping researchers refine phenotypic profiles and build translational bridges from animal models to human cognition.

Precision in behavioral neuroscience begins with observation—but its power is realized through technology. In a Morris Water Maze trial, each swim path tells a story, but it takes the right tools to capture, interpret, and quantify that narrative. When exploring advanced metrics like distance travelled in Whishaw’s Corridor, subtle differences in movement reflect crucial distinctions in learning, memory, and spatial navigation.
Modern behavioral tracking systems now make these distinctions measurable. High-frame-rate video capture paired with infrared lighting ensures accurate motion tracking even in opaque water. Real-time path plotting allows researchers to visualize each animal’s journey from the start location to the hidden platform, overlaid against digitally defined zones—including Whishaw’s Corridor itself.
Through automated software, swim trajectories are analyzed with centimeter-level precision. Researchers can customize corridor dimensions, track entry and exit points, and instantly calculate total distance travelled within that spatial window. These tools eliminate subjectivity and offer consistency across trials, researchers, and labs.
Advanced systems also support visual overlays like heatmaps and comparative traces. Researchers can monitor swim strategy evolution across time points, treatment conditions, or experimental groups—identifying, for example, whether an animal is shifting from procedural strategies to spatial ones. This capability makes it easier to assess the behavioral effects of neurological injury, pharmacological intervention, or genetic manipulation.
Data collection is paired with automated reporting, enabling seamless export of behavioral metrics and trajectory visuals for publication or collaboration. Cloud-based options ensure data can be accessed and reviewed across teams, accelerating data sharing while preserving integrity.
Trial structure is equally customizable—users can schedule randomized start positions, define probe days, or align analysis windows with treatment protocols or neural recording timepoints. These features allow researchers to extract precise, meaningful insights without sacrificing flexibility.
With a focus on spatially resolved data and intuitive analysis workflows, today’s behavioral platforms don’t just collect movement—they decode intention, strategy, and memory fidelity. When analyzing a rodent’s behavior within Whishaw’s Corridor, these systems bring clarity to the question: Did the animal remember, and how confidently did it act on that memory?
By combining environmental control, detailed motion tracking, and intelligent software, researchers are better equipped than ever to extract cognitive insights from navigation behavior—and do so with the rigor modern neuroscience demands.
In behavioral neuroscience, not all movement is equal. A rodent may reach the goal in a Morris Water Maze, but how it gets there—how directly, how confidently, how consistently—holds the deeper story. Distance travelled in Whishaw’s Corridor captures this story with exceptional granularity, offering a rare blend of cognitive specificity and spatial resolution. It’s a behavioral metric that moves beyond surface-level outcomes to interrogate the mechanisms of memory, decision-making, and neurological integrity.
Whether assessing hippocampal function in a control animal, detecting subtle spatial deficits in a disease model, or measuring therapeutic recovery after intervention, this metric provides a window into cognitive accuracy and behavioral certainty. It enables researchers to ask not just if an animal remembered, but how well, how efficiently, and how consistently that memory was expressed in action.
As research questions become more refined, and the demand for reproducible, translational science grows, so too does the need for analytical tools that maximize the meaning of every trial. Modern behavioral platforms now empower scientists to quantify movement in real time, define virtual corridors like Whishaw’s, and extract data that’s not just rich—but revealing.
This level of insight is now accessible and reproducible thanks to evolving technologies and experimental design frameworks that support customized zone mapping, automated trajectory analysis, and high-resolution tracking. These tools are reshaping how rodent behavior is interpreted—not just by measuring time or distance alone, but by recognizing intention, learning curve, and cognitive fidelity in every path taken.
As we continue to refine the metrics that define behavior, Whishaw’s Corridor reminds us that precision is more than a technical feature—it is a scientific ethic. The tools we use and the questions we ask must evolve together. For those committed to decoding the brain’s navigation of space, learning, and memory, the corridor offers a straight path to deeper understanding—and better science.
Written by researchers, for researchers — powered by Conduct Science.









Dr Louise Corscadden acts as Conduct Science’s Director of Science and Development and Academic Technology Transfer. Her background is in genetics, microbiology, neuroscience, and climate chemistry.




Monday – Friday
9 AM – 5 PM EST
DISCLAIMER: ConductScience and affiliate products are NOT designed for human consumption, testing, or clinical utilization. They are designed for pre-clinical utilization only. Customers purchasing apparatus for the purposes of scientific research or veterinary care affirm adherence to applicable regulatory bodies for the country in which their research or care is conducted.