Cellular Respiration Equation, Steps, Types and Importance

See our Environmental Science Products
What is Cellular Respiration?
Cellular respiration is a cellular catabolic process that transfers the energy embedded in carbohydrate molecules into energy carriers such as adenosine triphosphate (ATP).
Cellular respiration can take place with and without oxygen. Aerobic respiration occurs when oxygen is present, and it consists of four metabolic pathways: glycolysis, pyruvate oxidation, the Krebs cycle, and oxidative phosphorylation.
Without oxygen, it’s called anaerobic respiration, and it starts with glycolysis but ends when pyruvate is oxidized or decarboxylated. Carbon dioxide and water are the end-products of aerobic respiration.
Since ATP, including ADP, GDP, and phosphoryl group produced in cellular respiration, are reused in other pathways, the overall cellular respiration equation is expressed as the reverse of the photosynthesis equation:
C6H12O6 + 6O2 –> 6CO2 + 6H2O
Subsequently, the energy transferred to energy carriers supplies anabolic metabolism, which requires energy to synthesize macromolecules.
Summarily, as seen in the cellular respiration equation above, carbohydrates such as starch, glycogen, and sugars, are broken down, thus transferring the potential energy embedded in the chemical bonds to cellular energy carriers, which supply the energy to other anabolic pathways.

Figure 1: Structural Features of Mitochondria and the Overview of Aerobic Respiration
(Credit: “File:CellRespiration.svg” by RegisFrey is licensed under CC BY-SA 3.0. To view a copy of this license, visit https://creativecommons.org/licenses/by-sa/3.0)
Types of Cellular Respiration
1. Aerobic Respiration
Aerobic respiration occurs in most cells and organisms. It comprises four pathways: glycolysis, pyruvate decarboxylation, Krebs cycle, and oxidative phosphorylation (Figure 1).
In glycolysis, glucose molecules are broken down in the cell’s cytosol, resulting in the generation of pyruvate, ATP molecules, and the reduced form of nicotinamide adenine dinucleotide (NADH).
Pyruvate is subsequently transported to the mitochondrial matrix for decarboxylation, where acetyl coenzyme A (acetyl-CoA) is generated and enters the Krebs cycle.
As a result of the Krebs cycle, flavin adenine dinucleotide (FAD) and NAD+ are reduced and transported from the mitochondrial matrix to the mitochondrial inner membrane. Here, NADH and FADH2 participate in oxidative phosphorylation, where a series of electron transfers occur, with oxygen as the final electron acceptor.
Finally, the proton gradient across the mitochondrial inner membrane drives ATP synthesis in the matrix.
Generally, aerobic respiration is regarded as the “complete” cellular respiration since the overall biochemical reactions can be summarized as the reverse of photosynthesis.
2. Anaerobic Respiration
Typically referred to as fermentation, anaerobic respiration occurs when cellular oxygen is scarce or absent. It starts with glycolysis, where glucose is converted into pyruvate.
However, unlike aerobic respiration, the resulting pyruvate molecules are not transferred to the mitochondria. Instead, they remain in the cytosol and participate in one of the following reactions:
- Lactic Acid Fermentation
In lactic acid fermentation, pyruvate molecules from glycolysis accept electrons from NADH oxidation. The enzyme lactate dehydrogenase catalyzes this redox reaction:
CH3COCOO– + NADH + H+ <–> CH3CH(OH)COO– + NAD+
Pyruvate is reduced to lactic acid in this reaction, while NAD+ is regenerated to resupply glycolysis. Lactate can be used as a substrate in gluconeogenesis, which synthesizes glucose from non-carbohydrate substrates.
The given cellular respiration equation for glycolysis and lactate fermentation is summarized as follows:
C6H12O6 + 2ADP + 2Pi –> 2CH3CH(OH)COOH + 2ATP
Lactic acid fermentation occurs in prokaryotes and eukaryotes. Some prokaryotes are obligate anaerobes, which can only live in an oxygen-free environment. Others such as Lactobacillus, are facultative anaerobes; they’re predominantly anaerobic but are capable of aerobic respiration to a certain extent.[1-2] A third type known as aerotolerant anaerobes can tolerate the presence of oxygen but do not require it for growth.
- Ethanol Fermentation
Ethanol fermentation occurs after glycolysis and consists of two steps.
First, the enzyme pyruvate decarboxylase uses vitamin B1 as a coenzyme to cleave pyruvate, resulting in acetaldehyde and carbon dioxide. The reaction can be expressed as follows:
CH3COCOO– + H+ <–> CH3COH + CO2
Following the first reaction, acetaldehyde is reduced by NADH oxidation to NAD+. The enzyme alcohol dehydrogenase catalyzes this oxidation-reduction reaction in the following reaction:
CH3COH + NADH + H+ <–> CH3CH2OH + NAD+
Here, ethanol is produced along with the regeneration of NAD+. Like lactate fermentation, NAD+ is regenerated to resupply glycolysis, while ethanol is metabolized elsewhere.
The overall cellular respiration equation for glycolysis and ethanol fermentation pathway is as follows:
C6H12O6 + 2ADP + 2Pi –> 2CH3CH2OH + 2CO2 + 2ATP
Ethanol fermentation is typically found in eukaryotes such as yeast, plants, and certain vertebrate species. Moreover, most of these species are facultative aerobes, which predominantly use aerobic respiration but may use anaerobic respiration under certain circumstances.[1,3]
Both types of cellular respiration produce carbon dioxide, two ATP molecules and replenish the NAD+ pool. The energy gained is considerably less when pyruvate is transferred to the mitochondria for other glycolysis stages.
Nonetheless, anaerobic respiration is thought to be more flexible. It can replenish the cytosolic NAD+ pool and generate ATP considerably faster. The metabolites can also supply other metabolic pathways contributing to cellular energy transduction.
For this reason, anaerobic respiration in higher eukaryotes is thought to have evolved as a stress-response mechanism that inhibits or slows down glycolysis.
For example, strenuous muscle use in mammalian skeletal muscle cells results in lactate fermentation, which provides the cells with the necessary energy. It is also hypothesized that some plants maintain cellular osmotic potential by switching to ethanol fermentation when oxygen concentration is low.[3]
Four Stages in Cellular Respiration
1. Glycolysis
Considered the most critical pathway in carbohydrate metabolism, glycolysis occurs in both aerobic and anaerobic respiration. It can be split into two phases based on energy consumption, the investment and the payoff phases.
The investment phase uses ATP to split glucose molecules
When a glucose molecule enters glycolysis, it is initially converted to glucose-6-phosphate (G6P). The reaction is catalyzed by the enzyme hexokinase (or glucokinase for hepatocytes) and involves transferring a phosphoryl group from ATP to the glucose molecule. The reaction is irreversible; thus, it is regarded as the first committed step of glycolysis.
Afterward, G6P is converted to its isomer, fructose-6-phosphate (F6P), further transformed into fructose-1,6-bisphosphate (F1,6BP) phosphofructokinase. The conversion of F6P to F1,6BP is irreversible and requires ATP. Thus, it is regarded as the second committed step of glycolysis.
F1,6BP is split into glyceraldehyde-3-phosphate (G3P) and its isomer, dihydroxyacetone phosphate (DHAP), at the end of the investment phase. Both G3P and DHAP contain three-carbon molecules, and their structures are interchangeable by the enzyme triose phosphate isomerase.
Thus, the glycolysis investment phase can be added to the cellular respiration equation as follows:
C6H12O6 + 2ATP –> 2C3H5O6P-2 + 2ADP + 2H+
The payoff phase harvests energy during the generation of pyruvate
The payoff phase starts with the oxidation of G3P to 1,3-bisphosphoglycerate, coupled with reducing NAD+ to NADPH. The reaction is followed by the transfer of the compound’s phosphoryl group to ADP, generating the first ATP molecule from G3P along with 3-phosphoglycerate.
Afterward, 3-phosphoglycerate undergoes two subsequent rearrangement and bond cleavage reactions, resulting in phosphoenolpyruvate (PEP). Finally, PEP is converted to pyruvate when PEP’s phosphoryl group is transferred to ADP, generating the second ATP molecule from G3P.
Since G3P and its isomer, DHAP, are in equilibrium and interchangeable, DHAP is converted to G3P as the latter undergoes the reactions in the payoff phase. As a result, one glucose molecule, split into two G3P molecules during the investment phase, will give rise to four ATP molecules in the payoff phase.
This can be represented as:
C6H12O6 + 2ATP + 2ADP + 2Pi + 2NAD+ –> 2CH3COCOO + 4ATP + 2NADH + 2H+
Because two ATP molecules are consumed during the investment phase, the balanced equation is:
C6H12O6 + 2ADP + 2Pi + 2NAD+ –> 2CH3COCOO + 2ATP + 2NADH + 2H+
At the end of glycolysis, one six-carbon glucose molecule turns into two molecules of pyruvate and two NADPH molecules. It invests two ATP molecules in obtaining four ATP from substrate-level phosphorylation.
2. Pyruvate Oxidation
After glycolysis, pyruvate is reduced to replenish the cellular NAD+ reserve, depleted during glycolysis.
At this stage, cellular respiration can diverge to aerobic or anaerobic respiration. This is thought to be dictated by oxygen availability and the cellular NAD+/NADH ratio. When oxygen and NAD+/NADH ratio are low, pyruvate remains in the cytosol and undergoes anaerobic respiration.
When oxygen and cellular NAD+ are sufficient, pyruvate is sent to the mitochondria, transformed into acetyl-CoA.
Acetyl-CoA generation is achieved by the enzyme complex pyruvate dehydrogenase complex in the mitochondrial matrix. The complex consists of three enzymes, pyruvate dehydrogenase, dihydrolipoyl transacetylase, and dihydrolipoyl dehydrogenase. Each enzyme in the complex has its catalytic activity and requires coenzymes to function (Table 1).
They interact with pyruvate one after the other to catalyze its transformation into acetyl-CoA. Such coordinated action of an enzyme complex is termed substrate channeling.
The net reaction for pyruvate oxidation in aerobic respiration is:
CH3COCOO + CoASH + NAD+ –> CH3COCoA + CO2 + NADH + H+
When combined with glycolysis and pyruvate oxidation, the cellular respiration equation for pyruvate decarboxylation yields carbon dioxide, acetyl-CoA, and NADH:
C6H12O6 + 2ADP + 2Pi + 4NAD+ + 2CoASH –> 2CH3COCoA + 2CO2 + 2ATP + 4NADH + 4H+
Table 1: Enzymes in the pyruvate dehydrogenase complex and their corresponding coenzymes[1]
| Content | Pyruvate dehydrogenase | Thiamine pyrophosphate (TPP or vitamin B1) | Content | Dihydrolipoyl transacetylase | Lipoamide, coenzyme A (CoA) | Content | Dihydrolipoyl dehydrogenase | Flavin adenine dinucleotide (FAD), nicotinamide adenine dinucleotide (NAD+) |
3. The Krebs Cycle
Also known as the citric acid or tricarboxylic acid cycle, it is deemed the central pathway of aerobic respiration. It consists of a cycle of eight biochemical reactions in the mitochondrial matrix, which gives rise to precursors for the biosynthesis of amino acids and nucleotides and energy carriers in the form of ATP or guanosine triphosphate (GTP), NADH, and flavin adenine dinucleotide (FADH2).
Following pyruvate oxidation, a two-carbon molecule acetyl-CoA enters the pathways and reacts with oxaloacetate, a four-carbon molecule, in the presence of water. The reaction is catalyzed by citrate synthase and yields a six-carbon molecule, citric acid. It also releases coenzyme A (CoA) to the matrix.
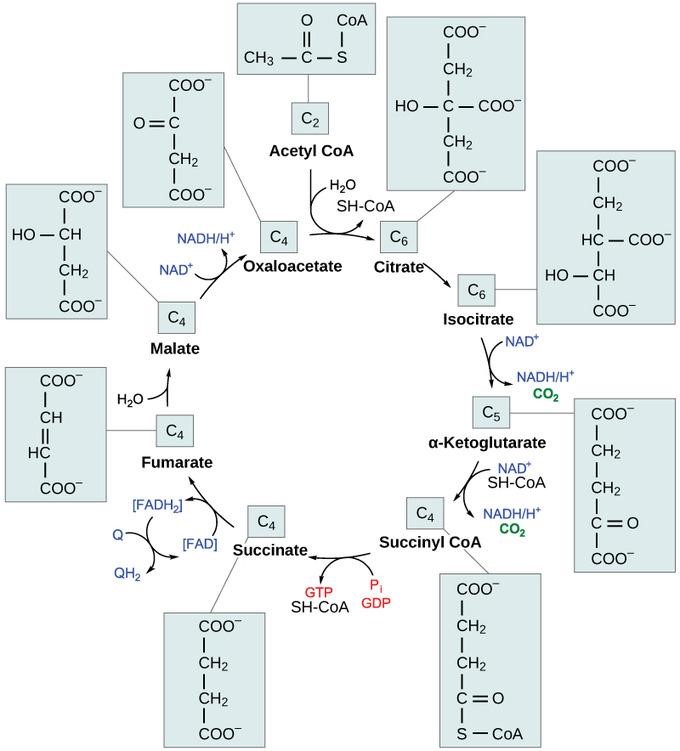
Figure 2: The Krebs cycle
Credit: courses.lumenlearning.com
In the second and third steps, citric acid undergoes a structural rearrangement reaction and oxidation before the six-carbon metabolite transforms into a five-carbon molecule, α-ketoglutarate, and releases carbon dioxide. The reaction is coupled with the reduction of NAD+ to NADH.
α-ketoglutarate is decarboxylated into four-carbon metabolites in the fourth and fifth steps. Succinyl-CoA is the first four-carbon metabolite regenerated with carbon dioxide and the NAD+ reduction.
Subsequently, the phosphoryl group in the coenzyme A is transferred from succinyl-CoA to ADP or GDP to generate succinate along with ATP or GTP from substrate-level phosphorylation.
To replenish oxaloacetate for the next round of the Krebs cycle, succinate is oxidized to fumarate by the enzyme succinate dehydrogenase, which requires its cofactor, FAD, to be simultaneously reduced to FADH2.
Water transforms fumarate into L-malate, which is eventually oxidized to oxaloacetate. This step is catalyzed by malate dehydrogenase and is coupled with reducing NAD+ to NADH.
The eight steps of the Krebs cycle are summarized as:
CH3COCoA + 3NAD+ + FAD + ADP or GDP + Pi + 2H2O –> 2CO2 + 3NADH + 3H+ + FADH2 + ATP or GTP + CoASH
Combining with the previous stages, the contribution of the Krebs cycle to the cellular respiration equation is:
C6H12O6 + 10NAD+ + 2FAD + 4ADP (or 2ADP and 2GDP) + 4Pi + 6H2O —> 6CO2 + 10NADH + 10H+ + 2FADH2 + 4ATP (or 2ATP and 2GTP)
As a whole, one molecule of acetyl Co-A that enters the Krebs cycle produces two carbon dioxide molecules. And since one glucose molecule is converted into two acetyl-CoA molecules, two rounds of the Krebs cycle generate four carbon dioxide molecules, six NADH, two FADH2, two ATP or GTP.
Like NADH generated in the previous stages, NADH and FADH2 donate electrons to the electron transport chain for oxidative phosphorylation. Carbon dioxide generated from cellular respiration is considered waste since it cannot resupply any other metabolic pathway.[1]
4. Electron Transport and Oxidative Phosphorylation
In the last stage of cellular respiration, the NADH and FADH2 produced from the previous stages are oxidized by the electron carriers, which are proteins in the electron transport chain (ETC).
Typically, the transfer of electrons during NADH and FADH2 oxidation occurs concurrently with the pumping of protons from the mitochondrial matrix across the inner membrane to the intermembrane space of the mitochondria.
The accumulation of protons in the mitochondrial intermembrane space establishes the proton gradients across the mitochondrial inner membrane. The proton gradient provides the energy for oxidative phosphorylation, which drives the phosphorylation of ADP by ATP synthase, synthesizing ATP in the mitochondrial matrix.
Electron transport sets up a proton gradient for oxidative phosphorylation
The ETC consists of several proteins that are electron carriers, which assemble into four complexes in the inner mitochondrial membrane:[4]
- Complex I: NADH-CoQ Oxidoreductase
Complex I of the ETC consists of the following electron carriers: flavin mononucleotide (FMN), an iron-sulfur cluster, and ubiquinone (UQ).
FMN is the first electron carrier that receives two electrons from NADH in the mitochondrial matrix, resupplying the mitochondrial NAD+ pool. The electrons from NADH are passed to the iron-sulfur cluster and eventually to UQ, reducing it to ubisemiquinone (UQH) and ubiquinol (UQH2).
Simultaneously, protons are pumped from the mitochondrial matrix to the intermembrane space, establishing the proton gradient that eventually drives ATP synthesis.
Also known as coenzyme Q (CoQ), UQ is the smallest and most hydrophobic electron carrier in the ETC. Thus, it only carries electrons on the membrane from one location to another.[1]
- Complex II: Succinate-CoQ Oxidoreductase
The second complex possesses the enzyme succinate dehydrogenase, which oxidizes succinate to fumarate and reduces FADH2 to FAD. The electrons donated from FADH2 are transferred to UQ, reducing the carrier to UQH2.[1,4]
Alternatively, the complex can contain acyl-CoA dehydrogenase instead of succinate dehydrogenase. In this case, the complex is linked to fatty acid catabolism called β-oxidation.[1]
The transfer of electrons in the second complex is not coupled with the pumping of protons across the mitochondrial inner membrane to the intermembrane space. Hence, it is the only complex in the ETC that does not contribute to the generation of proton gradients and the subsequent ATP synthesis.[4]
- Complex III: Cytochrome bc1 Oxidoreductase
Electrons are carried by UQH2 from Complex I or II to Complex III.
Complex III consists of cytochrome b andUQH2 carries electrons cytochrome c1 complexes, which contain an iron-sulfur protein component called the Rieske center that binds to UQH2 before passing the electrons to the heme prosthetic group cytochromes.
Because hemes can only accept one electron at a time, but UQH2 can carry two electrons simultaneously, the transfer of electrons to Complex III happens in a series of redox reactions called the Q cycle.
Briefly, when UQH2 arrives at Complex III, it binds to the Rieske center before it is oxidized, donating one electron to cytochrome c1. The electron in cytochrome c1 is sent to the cytochrome c at the peripheral of the complex, which carries the electron to Complex IV.
The reduction of cytochrome c1 results in an unstable UQH. Consequently, one electron in UQH is transferred to cytochrome b, reducing it and regenerating UQ. Following cytochrome b reduction, the electron is cycled back to regenerate the unstable UQH, further reduced to UQH2 from the electron donated from the second round of the Q cycle.
Similar to the transfer of electrons in Complex I, the Q cycle is coupled with the transfer of protons to the intermembrane space. Two protons are transferred when two electrons from two UQH2 reduce cytochrome c1. The other two protons are transferred from the matrix when each reduced cytochrome b donates its electron to regenerate UQ.
In total, two electrons from Complex III are carried to Complex IV via cytochrome c, and the other two are used to replenish UQH2. Hence, the Q cycle is thought to facilitate the transfer of electrons and maintain the proton gradients across the inner mitochondrial membrane.[1,4]
- Complex IV: Cytochrome c Oxidase
The last complex in the ETC comprises cytochrome a, cytochrome a3, a copper atom CuB, and a copper atom pair CuA center. They act as the redox center of the complex, which can accommodate four electrons.
At Complex IV, the electrons carried by four cytochrome c from Complex III are donated to the redox center. The electrons flow through the redox center until they are transferred to oxygen, which serves as the last electron acceptor of the ETC.
Since oxygen can accept four electrons, the flow of four electrons through the ETC will generate two water molecules. Simultaneously, four protons from the matrix are pumped across the inner membrane to the intermembrane space of the mitochondria.
Together, electron transport across complexes 1, 3, and 4 by the ETC can be explained as follows (this equation skips complex 2):
2NADH + 2H+ + O2 –> 2NAD+ + 2H2O
The electron transfer results in the regeneration of NAD+ and FAD in the matrix and the accumulation of protons in the intermembrane space of the mitochondria. The regenerated NAD+ and FAD can resupply other stages of cellular respiration in the mitochondrial matrix. The protons accumulated in the intermembrane space will eventually generate ATP during oxidative phosphorylation.[1,4]
ATP is produced by chemiosmosis in oxidative phosphorylation
The accumulation of protons in the intermembrane space of the mitochondria generates the proton motive force (or electrochemical gradient) that drives the synthesis of ATP. Unlike ATP synthesized in the previous stages of cellular respiration, ATP generated following electron transport in the inner mitochondrial membrane uses the chemiosmotic coupling mechanism.[1]
The chemiosmosis coupling mechanism involves the enzyme ATP synthase embedded in the inner membrane. Often referred to as Complex V of cellular respiration, ATP synthase is a unidirectional proton pump that transports the protons accumulated in the intermembrane space back to the matrix.
The transport of protons from the intermembrane space through ATP synthase leads to the collapse of the proton gradient across the mitochondrial membrane. This provides the potential energy that powers the phosphorylation of ADP, resulting in the synthesis of ATP in the mitochondrial matrix.
The two electrons from one NADH molecule flow through the ETC, starting from Complex I until they eventually reduce oxygen at Complex IV. Protons are pumped at Complexes I, III, and IV during electron transport, generating the proton gradient. It is estimated that one NADH, that is, two electrons, contributes to three ATP generation.
2NADH + 2H+ + O2 + 6ADP + 6Pi –> 2NAD+ + 2H2O + 6ATP
Similarly, the two electrons from one FADH2 molecule are donated to the oxygen at Complex IV. However, unlike NADH, FADH2 bypasses Complex I to Complex II, where FADH2 oxidation is not coupled with proton transfer. Therefore, FADH2 input to the proton gradient and the eventual ATP synthesis is less than that of NADH. It is estimated that one FADH2 molecule only generates 2 ATP molecules in the mitochondria.[1]
Thus, the oxidative phosphorylation equation is:
2NADH + 2H+ + 2FADH2 + 2O2 + 10ADP + 10Pi –> 2NAD+ + 2FAD + 4H2O + 10ATP
Based on the cellular respiration equation summarized until the Krebs cycle, one glucose molecule gives ten NADH and two FADH2. Hence, the cellular respiration equation at the oxidative phosphorylation stage is expressed as:
10NADH + 10H+ + 2FADH2 + 6O2 + 34ADP + 34Pi –> 10NAD+ + 2FAD + 14H2O + 34ATP
From glycolysis up to the Krebs cycle, cellular respiration has resulted in 4 ATP (or 2 ATP and 2 GTP), the cellular respiration equation with the contribution of oxidative phosphorylation is:
C6H12O6 + 6O2 + 38ADP (or 36ADP + 2GDP) + 38Pi –> 6CO2 + 6H2O + 38ATP (or 36ATP and 2GTP)
Importance and Relevance of Cellular Respiration
Deficiencies in any stage of cellular respiration can be consequential to the organism’s growth, well-being, and survival.
For instance,
- Mutations in genes encoding pyruvate carboxylase (PC) and succinate dehydrogenase (SDH) can disrupt glucose metabolism, including glycolysis and the Krebs cycle, and gluconeogenesis. PC and SDH deficiencies result in a broad spectrum of clinical symptoms, ranging from abnormal blood glucose concentration and urine profiles to developmental delay and metabolic acidosis.[5-6]
- OXPHOS diseases are clinical manifestations of mutations in the genes encoding proteins involved in oxidative phosphorylation. Neuro diseases such as Leigh syndrome, NARP (neurogenic muscle weakness, ataxia, and retinitis pigmentosa), and Deafness-dystonia-optic neuronopathy (DDON) syndrome are examples.[7-8]
In addition to the medical conditions due to deficiencies in cellular respiration, the understanding of cellular respiration is relevant to industrial applications.
For example,
- Lactate fermentation is the basis of food and beverage preservation and processing. Anaerobic respiration in lactic bacteria such as Lactobacillus and Streptococcus produces dairy products such as yogurt and cheese. For example, sourdough starters are lactic bacteria and yeasts metabolizing carbohydrate molecules (flour) during anaerobic respiration, producing carbon dioxide that lifts the dough and lactic acid that gives the dough its sour taste.[9]
- Ethanol fermentation in yeast is the main producing process of biofuels and other ethanol products such as alcoholic beverages or byproducts like spent yeast, protein concentrates, and industrial enzymes. Carbohydrate-rich materials such as honey, fruits, starchy grains, and tubers are fed to yeasts or other ethanol-producing microorganisms so that they can metabolize them during anaerobic respiration.[10]
In Conclusion
Cellular respiration is a catabolic process that harvests potential energy in carbohydrate molecules for other cellular activities. The process starts with the breakdown of glucose into pyruvate and diverges, depending on oxygen availability.
When oxygen is readily available, the cellular respiration equation reverses photosynthesis. In this scenario, pyruvate is sent to the mitochondria, where it is oxidized and fed to the Krebs cycle.
NADH and FADH2 are generated during these stages and later participate in the transport of electrons and oxidative phosphorylation, leading to the generation of ATP by chemiosmosis. Deficiencies in cellular respiration pose a tremendous effect on the health of organisms.
Anaerobic respiration occurs when oxygen is depleted or absent in the forms of lactate and ethanol fermentation. Either pathway shortens the cellular respiration equation since it does not produce ATP but quickly resupply glycolysis with NAD+. Both lactate and ethanol fermentation are relevant to the industrial production of food products, beverages, and biofuels.
References:
- Boyer R, Concepts in Biochemistry, 3rd edition. New Jersey: John Wiley & Sons; 2006.
- Zotta T, Parente E, Ricciardi A. Aerobic metabolism in the genus Lactobacillus: impact on stress response and potential applications in the food industry. J Appl Microbiol. 2017 Apr;122(4):857-869. doi: 10.1111/jam.13399.
- van Waarde, Aren. Alcoholic Fermentation in Multicellular Organisms. Physiological Zoology, 1991;64(4):895–920, http://www.jstor.org/stable/30157948.
- Voet D, Voet JG and Pratt CW, Fundamentals of Biochemistry, 2nd edition. New Jersey: John Wiley & Sons; 2006.
- Rustin, P., Munnich, A. & Rötig, A. Succinate dehydrogenase and human diseases: new insights into a well-known enzyme. Eur J Hum Genet 10, 289–291 (2002). https://doi.org/10.1038/sj.ejhg.5200793
- Wang D, De Vivo D. Pyruvate Carboxylase Deficiency. 2009 Jun 2 [Updated 2018 Mar 1]. In: Adam MP, Ardinger HH, Pagon RA, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993-2021. Available from: https://www.ncbi.nlm.nih.gov/books/NBK6852/
- Thorburn DR, Rahman J, Rahman S. Mitochondrial DNA-Associated Leigh Syndrome and NARP. 2003 Oct 30 [Updated 2017 Sep 28]. In: Adam MP, Ardinger HH, Pagon RA, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993-2021. Available from: https://www.ncbi.nlm.nih.gov/books/NBK1173/
- Benedicte O, Prete D, Mouni C. Mitochondrial Calcium Signalling: Role in Oxidative Phosphorylation Diseases. In: Bioenergetics. InTech; 2012. doi:10.5772/32117
- Petrova P, Petrov K. Lactic Acid Fermentation of Cereals and Pseudocereals: Ancient Nutritional Biotechnologies with Modern Applications. Nutrients. 2020 Apr 17;12(4):1118. doi: 10.3390/nu12041118. PMID: 32316499; PMCID: PMC7230154.
- Tse TJ, Wiens DJ, Chicilo F, Purdy SK, Reaney MJT. Value-Added Products from Ethanol Fermentation—A Review. Fermentation. 2021;7(4):267. doi:10.3390/fermentation7040267.