Photosynthesis is a series of biochemical reactions that photosynthetic autotrophs use to convert solar energy into potential energy, and it involves two stages.
In the first stage, called light-dependent reactions, autotrophs capture energy from sunlight. And in the second stage, light-independent or dark reactions, the captured solar energy is converted into potential energy, which exists in autotrophs as chemical bonds in macromolecules.
Most of the macromolecules synthesized from photosynthesis are carbohydrates, but autotrophs can also produce proteins and lipids. These macromolecules are consumed by the autotrophs in their metabolic reactions and utilized by heterotrophs as food and other non-food products.
Content
Chloroplast
A double-membrane organelle in photosynthetic eukaryotes such as plants where photosynthesis takes place.
Content
Granum (pl. grana)
A stack of a flattened sac-like structure called thylakoids in the chloroplast.
Content
Proplastid
An immature organelle found in eukaryotic autotrophs that contains no pigments, which matures and differentiates into chloroplasts and other plastids.
Content
Photon
An elementary particle or quantum of light which displays the properties of waves and particles. It is regarded as the smallest quantity of light.
Content
Phosphorylation
A chemical process of attaching a phosphate group in a cell. The inverse of the reaction is termed dephosphorylation.
Content
Photoreceptor
A light-sensitive protein complex that senses and responds to the presence of light.
Content
Photophosphorylation
A cellular chemical reaction that synthesizes ATP from the phosphorylation of ADP using solar energy.
Content
Thylakoid
A membrane-bound hollow disk-shaped structure in the chloroplast where photosynthesis in eukaryotes takes place.
Content
Stroma
A fluid that fills the inside of the chloroplast inner membrane.
What is Photosynthesis?
Photosynthesis occurs in autotrophs, which are organisms that are capable of producing their own food. They include archaea, prokaryotes such as purple bacteria, acidobacteria, and cyanobacteria, and eukaryotes such as algae and land plants.
Photosynthesis can be either anoxygenic or oxygenic. Anoxygenic photosynthesis does not require water or produce O2, and apart from cyanobacteria, photosynthesis in non-eukaryotes is anoxygenic.
Oxygenic photosynthesis is considered the reversal of glycolysis in cellular respiration. It captures energy from sunlight and uses it to convert CO2 into sugar and oxygen in the presence of water.
There are two stages of photosynthesis, which can be summarized as follows:[1,2]
(CO)2 + nH2O →(CH2O)n +O2;
where n represents the number of molecules.
In the first stage (light-dependent), the photoreceptors harvest energy from sunlight, and the harvested energy oxidizes water into oxygen (O2) and triggers the transfer of electrons. This leads to the reduction of nicotinamide adenine dinucleotide phosphate (NADP+) and the synthesis of adenosine triphosphate (ATP) from adenosine diphosphate (ADP) phosphorylation, in a reaction termed photophosphorylation.
The second stage, the light-independent reactions, are a part of the Calvin cycle. The reactions occur without the direct involvement of sunlight. Instead, the ATPs and NADPH generated in the light reactions are consumed to synthesize sugar from carbon dioxide (CO2). The resulting ADP and NADP+ are recycled to resupply the transfer of electrons in light-dependent reactions.[1]
Photosynthesis in Eukaryotes Occurs in Chloroplasts
Photosynthesis in eukaryotes takes place in chloroplast, an organelle genetically similar to cyanobacteria, leading to the endosymbiotic hypothesis which says that the origin of chloroplasts is comparable to that of mitochondria.
In particular, chloroplasts originated from heterotrophic eukaryotes that took up photosynthetic cyanobacteria. After endosymbiosis, the bacteria lost their ability to live independently, transforming the host eukaryotes from heterotrophs into autotrophs.[3]
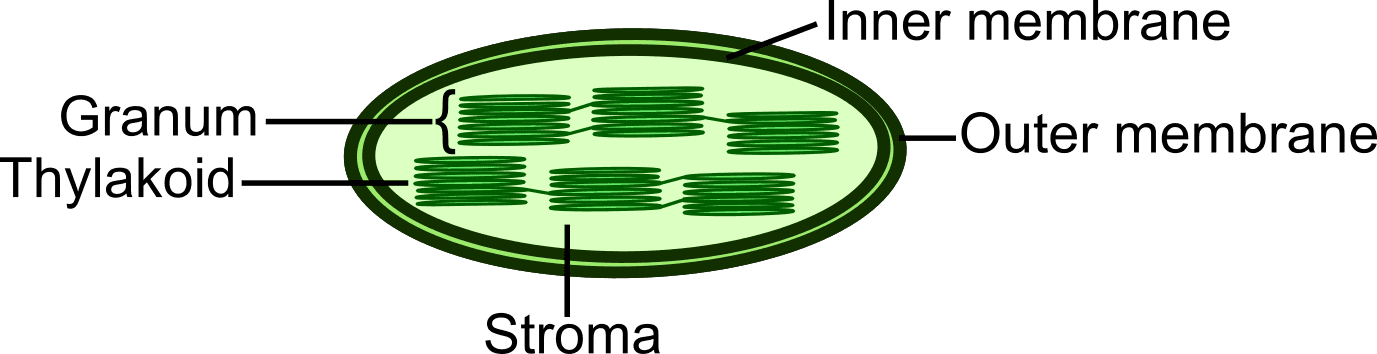
Figure 1: Structural Features of Chloroplasts in Photosynthetic Autotrophs
Similar to mitochondria, chloroplasts possess two layers of membranes, the outer and inner membrane. The outer membrane encloses the stroma, which contains several grana and other proteins. Each granum consists of a stack of thylakoids, a flattened sac-like structure, and the aqueous space in each thylakoid is called thylakoid lumen; however, the environment inside the thylakoid lumen is acidic (Figure 1).
Chloroplasts are present in cells and tissues involved in photosynthesis, for example, the guard cells and mesophylls in the leaf epidermis. They are differentiated from immature colorless plastids, called proplastids, and contain pigments that are predominantly chlorophylls, which give plants their green color.[1,2]
Photosynthetic pigments capture photons.
The sun radiates energy as electromagnetic waves, consisting of multiple wavelengths that inversely correspond to energy level. The smallest unit of energy carried by light is quantified in terms of light quanta orphotons. Photosynthesis in most autotrophs uses visible light, whose wavelengths range from 350 to 800 nanometers.[4]
Chlorophylls serve as the primary pigments that absorb light and capture photons in algae, cyanobacteria, and plants. They have a porphyrin ring consisting of four pyrroles that coordinate with one magnesium ion (Mg2+) in the center.
The ring has a long hydrocarbon side chain with one double carbon-carbon bond that enables chlorophylls to be solubilized in lipids and embedded in the thylakoid membrane.[1]
Two predominant chlorophyll species are found in plants and algae:[1]
Chlorophyll a is distinguished by a methyl group attached to one of the four pyrrole rings. It is the most prevalent chlorophyll species in plants and the only one present in the photosynthetic reaction centers. Chlorophyll a absorbs the most light at 372 and 642 nanometers, corresponding to violet-blue and orange lights, respectively.[1,4]
Chlorophyll b is characterized by a formyl group attached to the porphyrin ring in the same position as the methyl group in chlorophyll a. The substitution of the formyl group shifts the optimal light-absorbing ranges in chlorophyll b to 392 and 626 nanometers, corresponding to blue and red lights, respectively.[1,4]
Apart from chlorophylls, other accessory pigments are also present in photosynthetic cells. These accessory pigments absorb visible light of different wavelengths, supporting the energy capturing process. Accessory pigments can prevent intense or excessive light from damaging chlorophylls and photosynthetic components by reducing free radicals.[1, 2]
Examples of accessory pigments are:[1,2]
Carotenoids are pigments that absorb blue-green and violet lights, corresponding to 400 to 500 nanometers. They give algae and plants yellow, orange, and red colors. Examples are carotenes which are unsaturated hydrocarbons carotenoids, and xanthophylls which are oxygenated carotenoids.
Phycobilins are red pigments that absorb lights in the 550 to 630 nanometers range. Unlike carotenoids, phycobilins are found only in red algae and some unicellular algae (like cryptomonads) but not in plants or green algae.
Since all photosynthetic pigments absorb light at different wavelengths, the combined light-absorbing action of all pigments will allow organisms to maximize the capturing of photons.[1]
Mechanisms of Photosynthesis
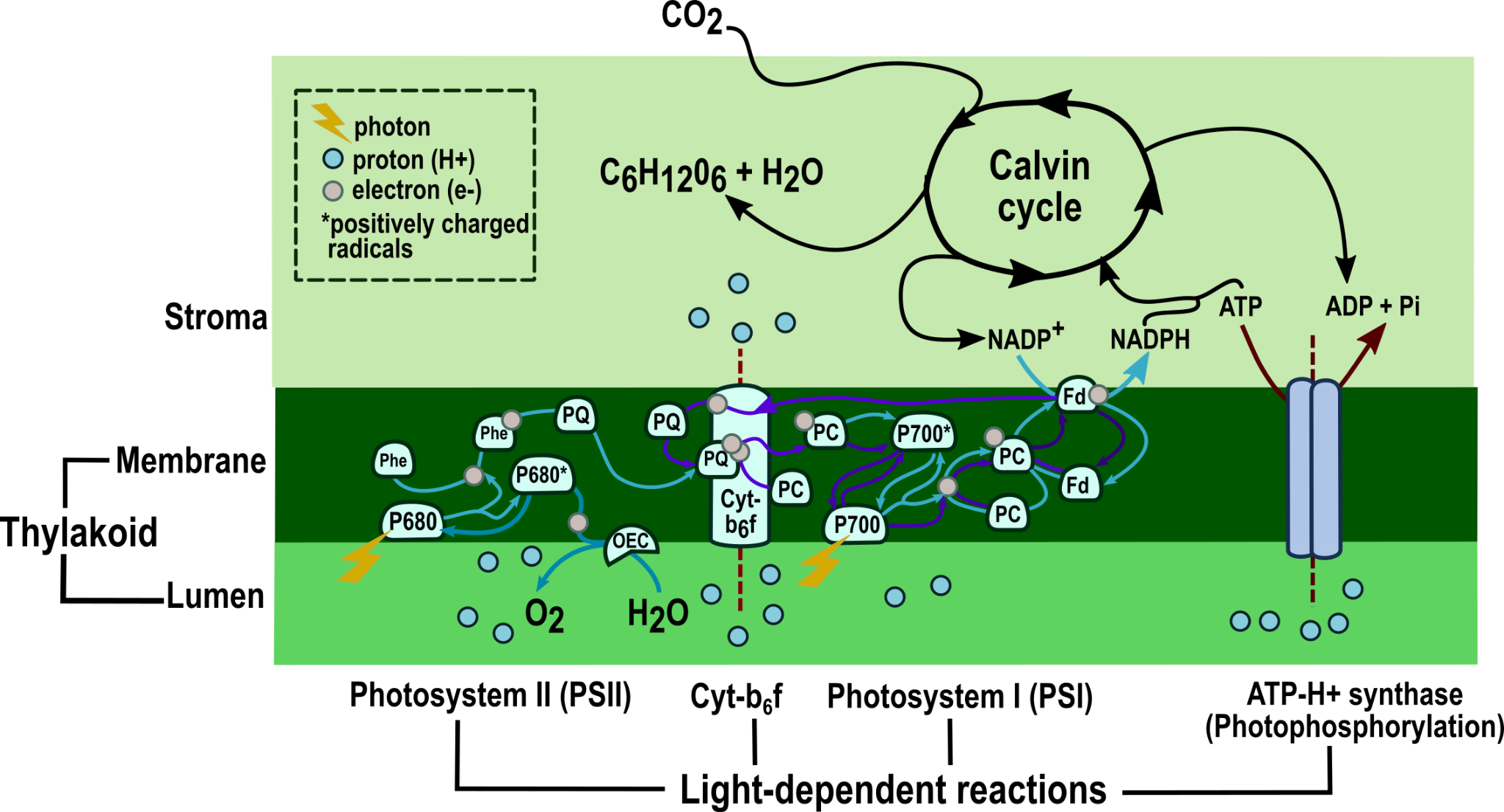
Photosynthesis mechanisms can be divided into two stages based on the light requirement. The two stages of photosynthesis are connected by two high-energy molecules, ATP and NADPH (Figure 2). Both ATP and NADPH are consumed in the second stage when hexose sugar is synthesized. Hexose sugars are building blocks for the synthesis of complex carbohydrates.
Figure 2: Mechanism of photosynthesis — Summary of its two stages: light-dependent reactions and Calvin cycle. The first stage consists of light harvesting, electron transfer and photophosphorylation. Photons are harvested from light by photoreceptors in the photosystems I and II, triggering the transfer of electrons between the two photosystems and the protein complex, cytochrome b6f. Electron transfer generates proton gradients, which drive photophosphorylation, as depicted by dark red arrows. Non-cyclic photophosphorylation, known as the Z scheme, results from the transfer of electrons shown by light-blue arrows, and cyclic photophosphorylation results from the transfer of electrons represented by purple arrows. The light reactions produce ATP and NADPH consumed in the Calvin cycle when hexose sugar (C6H12O6) is generated.
Credit: (Modified from Boyer, 2006 and Heldt, 2005).[1, 2]
A. Light-dependent Reactions
As indicated in the name, photosynthetic light-dependent reactions occur when light is present. In eukaryotes, the light-dependent reactions take place in the thylakoid membrane in three steps (Figure 2):[1]
1. Harvesting of Photons from Light
In this step, chlorophylls, accessory pigments, and chlorophyll-associated proteins act as photoreceptors to harvest photons from light. Photoreceptors are assembled into functional groups consisting of a photosynthetic reaction center, light-harvesting (LHC), and core antenna complexes.
The accessory pigments and chlorophyll-bound proteins make up the core antenna and LHC complexes. They support the photosynthetic reaction center by acting as antennae that capture photons from lights of various wavelengths. The captured photon is transferred one by one until it reaches the photosynthetic reaction center.
Plants, green algae, and cyanobacteria possess two photosynthetic reaction centers. They are arranged in tandem and connected via the cytochrome b6f complex.
The first photosynthetic reaction center, photosystem I (PSI), uses chlorophyll a as the primary photoreceptor. It can be excited by light at the maximum wavelength of 700 nanometers (P700).[2, 5]
The second reaction center, photosystem II (PSII), uses chlorophyll a and b to absorb photon energy. PSII can absorb photons from light up to the wavelength of 680 nanometers (P680).[2, 5]
2. Electron Transfer
Harvesting photons results in photoinduced charge separation. It excites an electron of the photoreceptor to a higher energy level, forming a negatively charged radical.
The instability of the excited state precipitates in spontaneous electron transfer to a nearby acceptor molecule, which leaves the photoreceptor positively charged. Eventually, the photoreceptors are replenished with electrons from water or another electron transfer chain.[1-2, 5]
The transfer of electrons occurs in the photosystems and cytochrome b6f complex (Cyt-b6f) as follows:[1, 2, 5]
When the harvested photon reaches the reaction center (P700), the excited electron is transferred to plastocyanin (PC).
Afterwards, the oxidized PC donates the electron to the next available acceptor. Once the electron reaches the stroma side of the thylakoid membrane, it oxidizes ferredoxin (Fd), while NADP+ in the stroma is simultaneously reduced to NADPH.
NADPH produced in PSI is subsequently used in carbohydrate synthesis. The -positively charged P700 is resupplied with electrons transferred from Cyt-b6f.
PSI (Plastocyanin-Ferredoxin Oxidoreductase)
At PSII, the harvested photon excites the P680, igniting an electron transfer. The negatively charged P680 sequentially donates electrons to pheophytin (Phe) and plastoquinone (PQ). Upon accepting electrons, PQ is reduced to plastosemiquinone (PQH) and plastoquinol (PQH2), which subsequently diffuses to Cyt-b6f.
The positively charged P680 is replenished with electrons from water. The water-splitting complex, also called the oxygen-evolving complex (OEC), catalyzes this reaction.
OEC is a metalloenzyme located on the luminal side of the thylakoid membrane. It contains manganese ions in its catalytic center and uses them as cofactors to initiate the oxidation of water. As a result, O2 is generated and released to the atmosphere, the electrons cancel the positively charged P680, and the residual protons (H+) are accumulated in the thylakoid lumen.
Cytochrome b6f complex (Cyt-b6f) -connects the transfer of electrons between the photosystems. As a result, the flow of electrons between PSII and PSI is linear and follows the so-called Z scheme.
Here, plastoquinol (PQH2) carries the electrons from PSII to Cyt-b6f. PQH2oxidation takes place sequentially, and electrons are eventually transferred to plastocyanin (PC). The resulting PQ refills the PSII, while the reduced PC carries the electron to replenish the positively charged PS700 in the PSI.
Since PQH2 is generated in the stroma-facing side of the thylakoid in PSII, PQ oxidation in Cyt-b6f involves the transfer of electrons to the luminal side of the thylakoid membrane. Thus, electron transfer in Cyt-b6f is coupled with proton transfer across the thylakoid membrane. This leads to the generation of proton gradients, which provide energy for photophosphorylation.
3. Photophosphorylation[1, 2]
Z scheme pathway (non-cyclic photophosphorylation)
The transfer of electrons after light-harvesting generates a proton gradient across the thylakoid membrane.
Protons are deposited in the thylakoid lumen in the Z scheme during water and plastoquinol (PQH2) oxidation reactions. The accumulated protons generate the electrochemical potential that drives ATP synthesis by ADP phosphorylation via ATP-H+ synthase.
Alternative to the Z scheme, ATP can be synthesized without reducing NADP+ in cyclic electron flow.
Here, the electron flow is restricted between PSI and Cyt-b6f. The excited electron in the P700 is transferred to PC and ferredoxin (Fd). However, the oxidized Fd does not donate electrons to NADP+. Instead, oxidized Fd is directed to Cyt-b6f, where the electrons are donated to plastoquinol and plastocyanin, respectively, while the reduced PC donates its electron to recharge the PS700.
Like the Z scheme, electron flow in Cyt-b6f is coupled with proton transfer, generating the proton gradient across the thylakoid membrane that drives the conversion of ADP to ATP.
B. The Calvin(-Benson-Bassham) Cycle
The second stage of photosynthesis requires NADPH and ATP from the light-dependent reactions for several reactions in the Calvin cycle.
The Calvin cycle is the reductive pentose phosphate pathway based on the reaction nature and the reactant (pentose). The reactions in the cycle occur in the stroma without the direct involvement of light. Hence, the nickname dark reactions.
The cycle consists of the following stages:[2]
1. Carbon Dioxide Assimilation
Atmospheric CO2 is fixed and incorporated into ribulose-1,5-bisphosphate (RuBP), a five-carbon sugar molecule. The incorporation results in the formation of two 3-phosphoglycerate molecules.
The assimilation of CO2 to RuBP is highly exergonic, suggesting that it is nearly irreversible. The reaction is catalyzed by the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO). RubisCO is the only enzyme known to date that is capable of fixing CO2 from the atmosphere. It is also the most abundant enzyme in plants and one of the largest.[2]
2. Reductive Pentose Synthesis
The two molecules of 3-phosphoglycerate generated from CO2 assimilation undergo successive reductive reactions, transforming them into a three-carbon glyceraldehyde-3-phosphate (G3P) and its isomer, dihydroxyacetone phosphate (DHAP). These reactions consume the ATP and NADPH generated in the light reactions.[1-2]
At this point, one-sixth of G3P and DHAP molecules are condensed to form a six-carbon phosphate sugar molecule, fructose-1,6-bisphosphate (FbisP). Later, FbisP is irreversibly hydrolyzed into fructose-6-phosphate, serving as a reactant in starch and cellulose biosynthesis or converted to glucose.[1] The majority of G3P and DHAP are used to regenerate RuBP.[3]
3. Regeneration of Carbon Acceptor Molecules
In the last stage, two DHAP and three G3P molecules go through a series of enzymatic reactions to regenerate three pentose phosphate molecules.
Each pentose phosphate contains five carbon molecules and a phosphate group. ATP phosphorylates all three pentose phosphates to become RuBP for the next round of the Calvin cycle.
The reactions in the Calvin cycle can be summarized as follows:[1]
Based on the summary, it can be roughly estimated that the fixation of one CO2 molecule consumes two NADPH molecules and three ATP molecules.
Photorespiration is the Calvin Cycle Bad Company
RubisCO possesses both carboxylase and oxygenase activities. For this reason, the enzyme can fix O2 and incorporate it into RuBP. Consequently, a two-carbon compound, phosphoglycolate and three-carbon compound, 3-phosphoglycerate (3PGA), are generated instead of two 3-phosphoglycerate molecules.
The generated 3PGA can enter the Calvin cycle and synthesise hexose sugar, while the two-carbon metabolite is first transported to peroxisomes for modification. The modified metabolite is transported to mitochondria and decarboxylated, releasing one molecule of CO2. Then, the metabolite is transported back to the peroxisome and converted into glycerate, which can be converted into 3PGA that can enter the Calvin cycle.[2]
The conversion of phosphoglycolate into 3PGA is regarded as a wasteful process. Not only CO2 is lost, but ATP and NADPH are also consumed in the process. Nevertheless, it is thought that photorespiration could prevent photodamage by consuming surplus ATP and NADPH.[2]
To circumvent photorespiration, alternative CO2 assimilation methods have evolved to reduce RubisCO activity. For instance, C4 and CAM plants use the enzyme phosphoenolpyruvate carboxylase, or PEP carboxylase, to capture CO2 in specialized cells.[1]
Factors Influencing Photosynthesis
The rate of photosynthesis depends on the efficiency and the effectiveness of its two stages. Based on its mechanisms, several environmental and internal factors influence the two stages of photosynthesis, including:
1. Light Quality and Quantity
Light can initiate photosynthesis only when photoreceptors are excited by the photons. Since photoreceptors only perceive light of limited ranges, not all light will contribute to photosynthesis, and the capacity to initiate photosynthesis is also different for each wavelength.
In addition, photoreceptors are proteins that can withstand only a certain level of energy before they are permanently denatured. For example, a slight increase in the light intensity, i.e. the number of photons, could enhance the rate of photosynthesis.
However, a significant increase in light intensity can cause photodamage that destroys photoreceptors and other photosynthetic machinery. Excess photons can also trigger photoinhibition, causing disruptions in the transfer of electrons at PSII.
2. Water Availability
Water is a prerequisite for the light-dependent reactions and Calvin cycle. Thus, a decrease in water will affect the production of ATP and NADPH in the light-dependent reactions and synthesis of hexose sugar in the Calvin cycle.
Water deficiency could delay or disrupt the electron flow in the PSII because water serves as electron acceptors that replenish the charged P680. Similarly, reactions in the Calvin cycle are also disturbed due to the decrease in the amount of water that can participate or the reduction of ATP and NADPH available from the light reactions.
3. Carbon Dioxide Concentration
As one of the reactants of the Calvin cycle, the increase in atmospheric CO2 can accelerate the synthesis of carbohydrates and reduce photorespiration.
Nonetheless, CO2 is fixed at the guard cell stomata, where transpiration also takes place. In other words, CO2 fixation occurs at the expense of water availability, limiting the extent to which CO2 can stimulate the rate of photosynthesis.
4. Temperature
Temperature can act on photosynthetic machinery such as photoreceptors and enzymes in the Calvin cycle. Their 3-dimensional structures and catalytic activities slightly differ when the temperature shifts. However, extreme changes in the temperature on either end can significantly alter both the structure and activity of photosynthetic machinery.
Moreover, temperature change can also affect the transpiration rate, implying a change in water availability. As previously discussed, water plays a direct role in the photosynthetic electron transfer, influencing the synthesis of ATP, NADPH and carbohydrates.
5. Genetic Predisposition
Apart from environmental factors, the efficiency of photosynthesis also depends on the internal factors of the organisms of interest. These factors are genetic predispositions, which dictate the working of the biological systems.
For example, genetic variations in the photoreceptors and related enzymes can be translated into differences in their functions and catalytic abilities. On a larger scale, some green algae and plants species have evolved new mechanisms that minimize wasteful processes and enhance the efficiency of photosynthesis.
In Conclusion
All in all, photosynthesis is a complex biochemical process that captures solar energy for food production. The two stages of photosynthesis include the light-dependent reactions where photoreceptors harvest photons, triggering a series of electron transport. The first stage releases oxygen to the atmosphere and generates ATP and NADPH for the second stage.
Carbohydrates and other macromolecules are synthesized from ATP and NADPH-consuming reactions in the Calvin cycle (second stage). These macromolecules not only feed the autotrophs but are also consumed and used by other organisms in the ecosystem.
Thus, photosynthesis is a process that provides the organisms in the ecosystem with the air to breathe, food to eat, and raw materials for manufacturing non-food products.
References:
Boyer R, Concepts in Biochemistry, 3rd edition. New Jersey: John Wiley & Sons; 2006.
Heldt H-Wa. Plant Biochemistry. 3rd ed. San Diago, California: Academic Press; 2005.
Martin WF, Garg S, Zimorski V. Endosymbiotic theories for eukaryote origin. Philos Trans R Soc B Biol Sci. 2015;370(1678):20140330. doi:10.1098/rstb.2014.0330
Milne BF, Toker Y, Rubio A, Nielsen SB. Unraveling the Intrinsic Color of Chlorophyll. Angew Chemie Int Ed. 2015;54(7):2170-2173. doi:10.1002/anie.201410899
Gao J, Wang H, Yuan Q, Feng Y. Structure and Function of the Photosystem Supercomplexes. Front Plant Sci. 2018;9. doi:10.3389/fpls.2018.00357