
Scientific Equipment for Every Lab
From behavioral mazes to stereotaxic instruments, lab consumables to analytical tools. Trusted by 1,200+ research institutions worldwide.
Browse by research area.







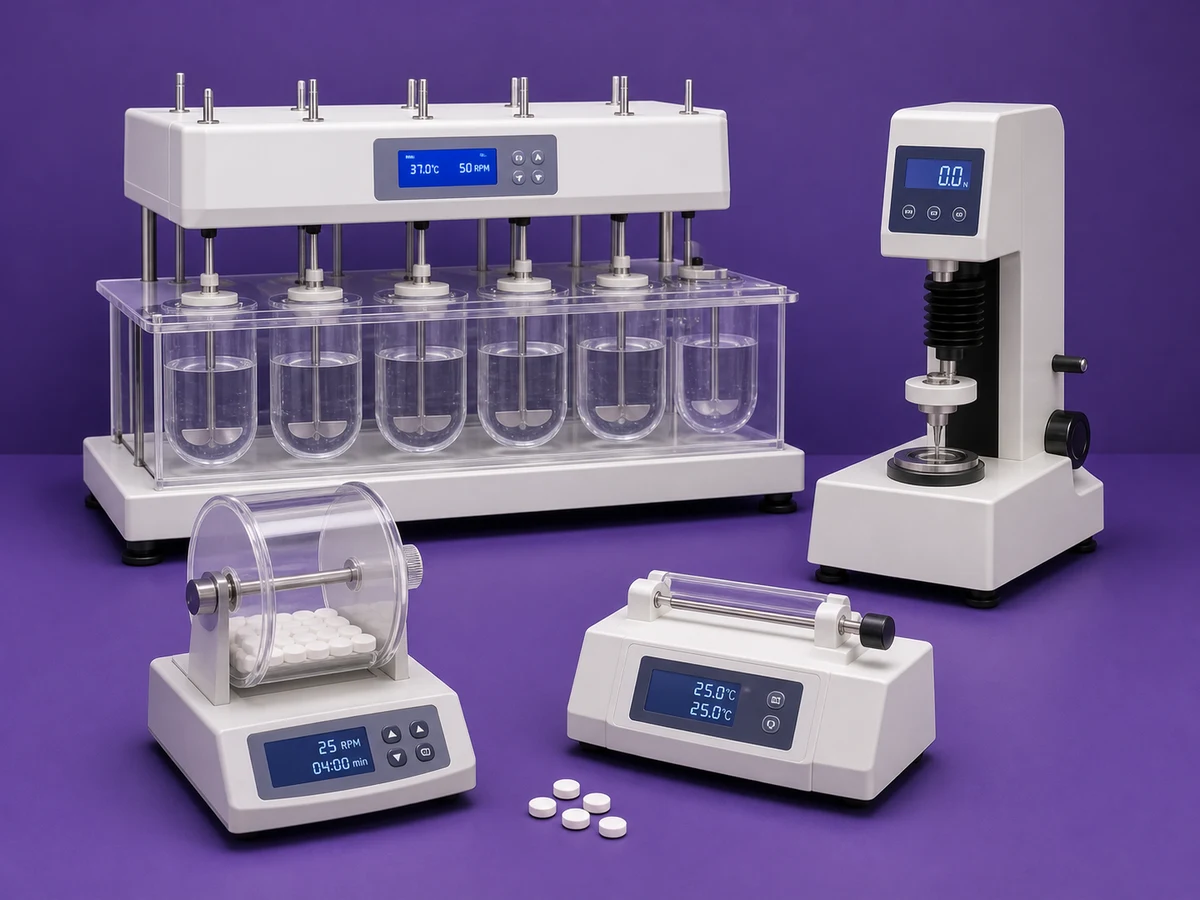
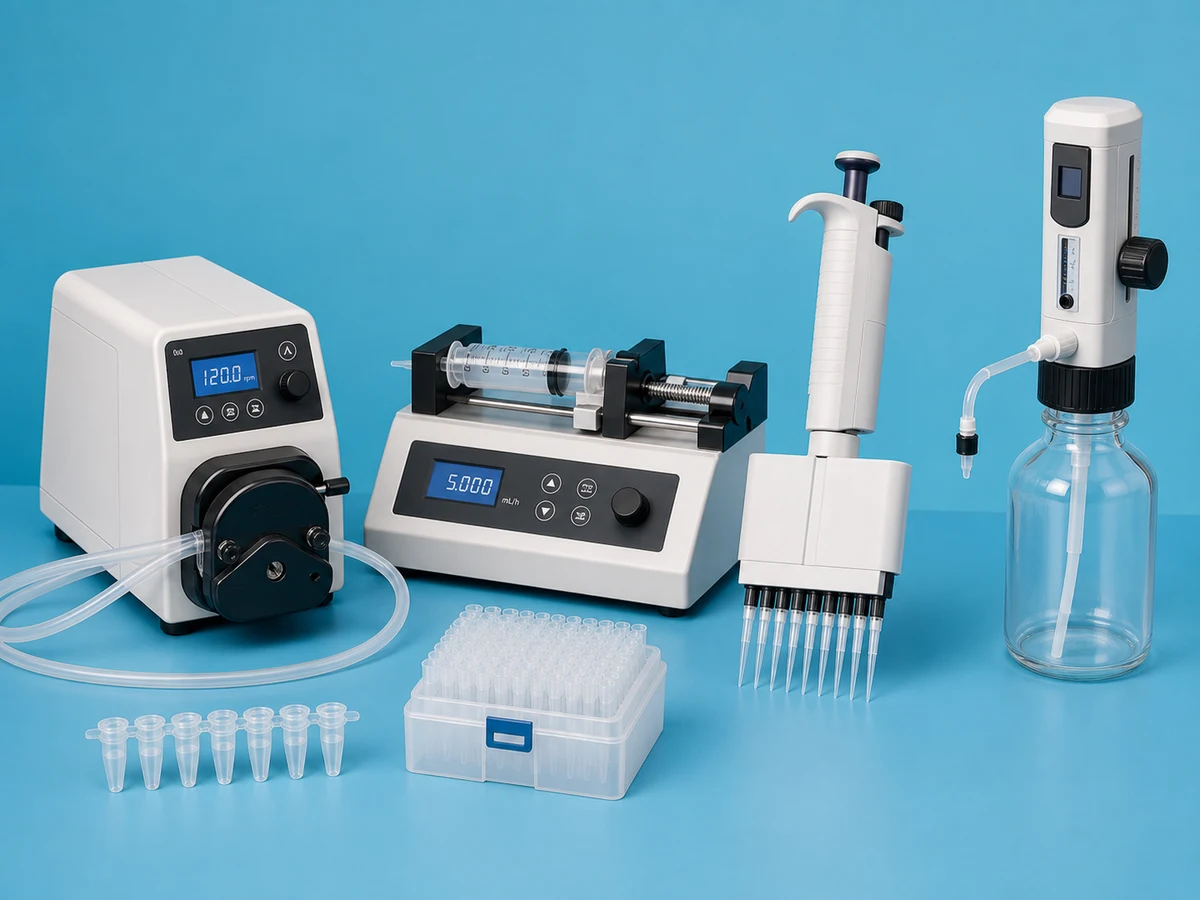

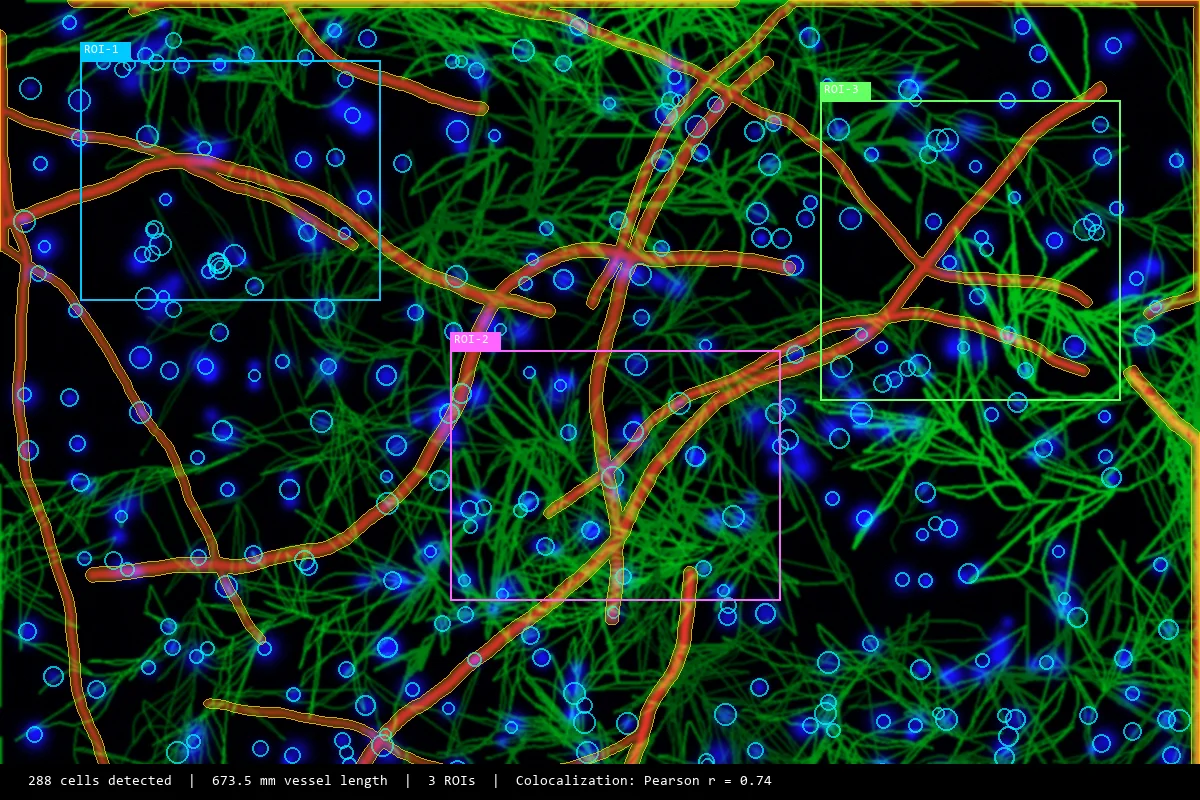
High-accuracy behavioral analysis for every experiment.
ConductVision tracks rodent, zebrafish, and drosophila activity in real time. 95–99% pose precision, 10+ species, runs locally on the desktop hardware we ship.



Trusted by world-class researchers.
“Got two open-field mazes as well as a camera gantry to run some behavior testing. Customer service was quick to respond to our questions and provide clear answers. The quality of the gantry and open field mazes seems to be excellent.”
World-class behavioral mazes. Find yours.
From the open-field arena to the most complex decision-making paradigms. Designed for reproducibility, shipped to spec.






Software for every step between the question and the paper.
Plan the study. Manage the colony. Score the behavior. Run the cognition test. File the IRB. Each tool reads and writes the same study record, so the lab can publish without rebuilding the dataset.
All softwareInstitutional procurement.
Standing orders, institutional terms, and the parts of the catalog procurement actually owns — not the bench-by-bench browse above.
 Bulk
BulkConsumables & PPE
Gloves, tubes, parafilm, lab coats, eyewear. Bulk pricing on recurring spend.
 Build
BuildLab Furniture & Build-out
Benches, cabinets, sinks, carts. Whole-room outfitting and CAPEX projects.
CAPEXCAPEX Quote Packages
Multi-item quote packets with specs, freight, install, warranty, and lead times in one place.
 Account
AccountMSA & Preferred Vendor
Pre-negotiated pricing for institutions on master service agreements.
 Auto-ship
Auto-shipRecurring Orders
Standing weekly or monthly deliveries — set once, forget.
 Setup
SetupInstitutional Accounts
Net-30 terms, W-9, BAA, vendor onboarding — handled by our procurement team.
Knowledge, before the order.
The Science Library.
Open methodology references built up over a decade. Protocols, paradigms, surgical guides, histology methods, and equipment reviews — every page links back to the products that run them.
Searchable directory of every paper-citing equipment and method we support, indexed by paradigm and species.
Sample-size calculators, dose converters, freezer selectors, lab planners — free, no sign-in.
Standalone Harvard-style CV builder for promotion packets and grant applications.
In-house studies, replication notes, and methodology reviews.
Executable, version-tracked behavior protocols you can hand to a tech.
Sister registry: per-experiment records that travel with the paper.
Complete walkthroughs to stand up behavioral, histology, surgery, and cell-culture labs.
Reference of every behavioral paradigm we support — parameters, citations, equipment.
Built around the science.
Custom Configurations
Every lab is different. We customize equipment dimensions, materials, and specifications to match your exact research protocol.
Fast Quote Turnaround
Average 6-day quote-to-order cycle. Get pricing within 24 hours for standard equipment, 48 hours for custom builds.
Trusted by Academia
From NIH and Calico to Cornell and Emory, over 1,200 institutions trust ConductScience for their research equipment.
24/7 Technical Support
Expert support whenever you need it. Our team includes scientists who understand your research, not just the equipment.