Biological Membrane: Definition, Structure, Components, and Functions

Introduction and History
The biological membrane, also known as cell membrane or biomembrane, is a selectively permeable membrane that defines the boundary of the cell and separates it from the external environment.[1]
The existence of cell membranes wasn’t known until the invention of the microscope in the seventeenth century.[2] With microscopic observations, scientists came to know that organisms are composed of cells but the existence of a biological membrane was still out of the equation.
It took around 200 years (in the 19th century) to learn that there’s some form of a semi-permeable barrier separating the cell from its surrounding environment.[2] But, the composition of the biological membrane was not known until the discovery of electron microscope in the 1950s.[2]
The meticulous work and findings of several scientists like Overton, Langmuir, Gorter, and Grendel, and Davson and Danielli deduced the presence of lipids and proteins in the biological membrane.[3] The modern concept of lipid bilayer was proposed late in 1972, after a tough debate on the arrangements of these molecules in the cell membrane.[2]
This article brings you all about biological membranes, from their different models proposed by scientists, structural organization and composition, to their functions in different organisms.
Biological Membrane Models
Understanding all proposed membrane models in the past led to the current concept and characteristics of the fluid mosaic model. Some of the popular membrane models proposed by scientists and their significance are given below.
1. Gorter and Grendel’s membrane theory (1920)
Evert Gorter and François Grendel hypothesized that if the plasma membrane is a bi-layer, then the surface area of the monolayer of lipids measured would be double the surface area of the plasma membrane.[3]
They supported their assumption by using the blood cells of a few mammals and measuring the total surface area of the plasma membrane of red blood cells and the area of the monolayer of lipids using Langmuir’s method.[3]
They proposed the plasma membrane structure as a lipid bilayer with the hydrophilic head of the lipid on the outside of the membrane and hydrophobic tail facing away from the aqueous environment.[3]
Apart from their proposed structure, the scientists couldn’t completely support their hypothesis. The model assumed a higher population of lipids in the membrane with no supporting ground and couldn’t describe the properties and functions of the membrane.[3]
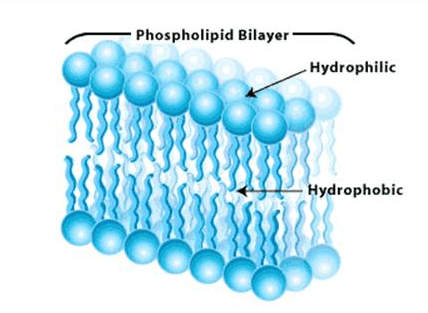
Image: Gorter and Grendel’s cell membrane model.[4]
Source: Biology4isc
2. The Davson and Danielli Sandwich Model (1935)
Davson and Danielli proposed that the cell membrane is composed of a lipid bilayer with a thin layer of protein coating on both sides of the membrane.[3] They explained that the cell membrane contains a lipoid center covered by protein monolayers.
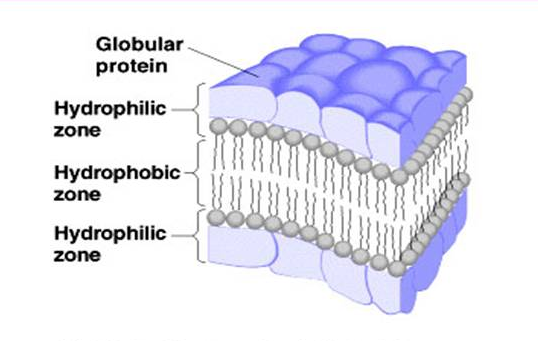
Image: An illustration of the sandwich model of the cell membrane as proposed by Davson and Danielli.[3]
Source: Biology4isc
3. Robertson’s Unit Membrane Model (1950)
Robertson’s model supported Davson and Danielli’s sandwich model. Using high-resolution electron microscopy, he observed a trilaminar pattern of the cell membrane.[3]
From this observation, he deduced that all cellular membranes share a similar underlying structure, the unit membrane, and the membranes consist of a lipid bi-layer covered on both surfaces with thin sheets of proteins.[3]
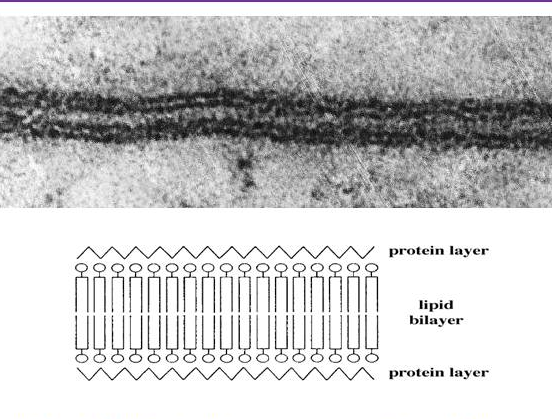
4. Singer and Nicolson’s fluid mosaic model (1972)
This is the most accepted model among all the proposed models. It says that the cell membrane is composed of a lipid bilayer with proteins embedded in the membrane.
It proposes three types of proteins in the membrane in a mosaic pattern: integral protein, peripheral proteins, and lipid anchored proteins.
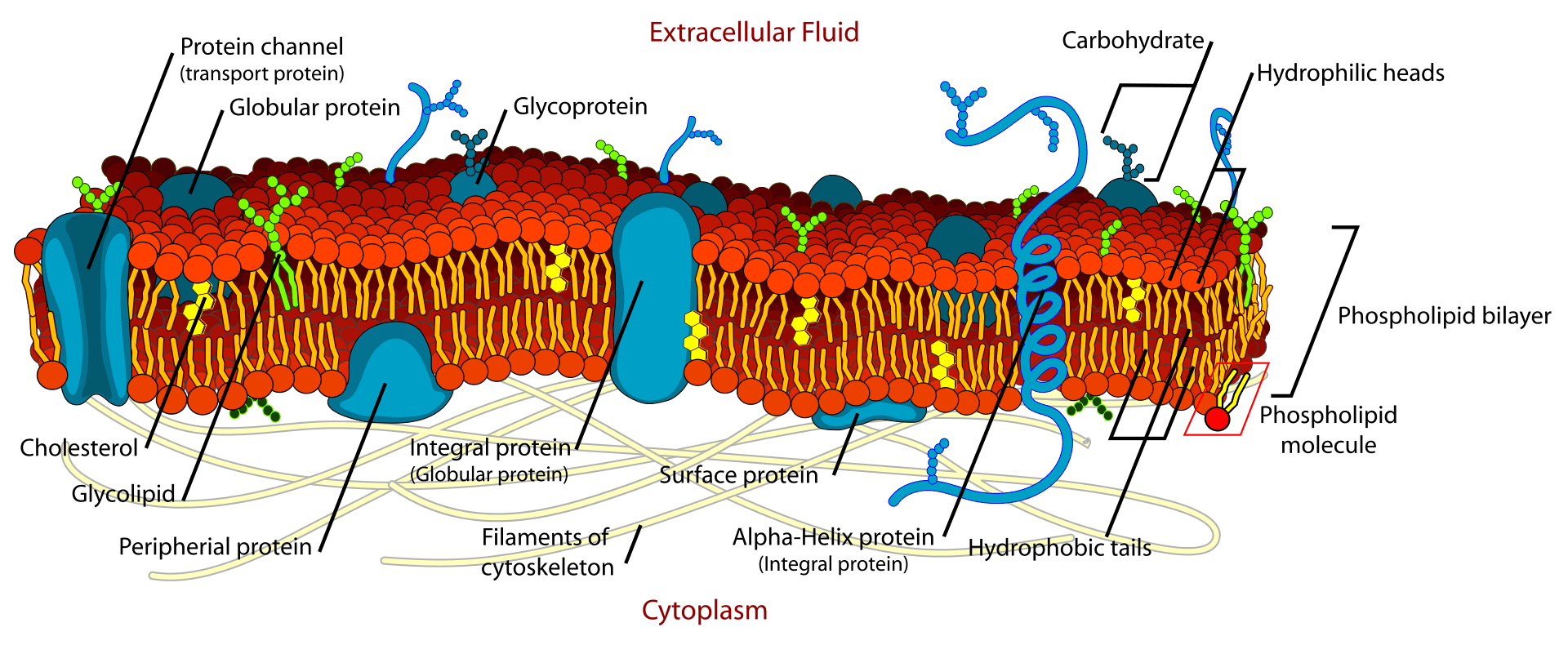
Image: An illustration of the fluid mosaic model proposed by Singer and Nicolson.[3]
Source: Wikipedia
{kind=link}
Structure and Components of the Cell Membrane
As the fluid mosaic model proposes, the cellular membrane is composed of both lipids and proteins. All membrane structures share a common organization: a phospholipid bilayer with embedded proteins.[5] The membrane proteins are responsible for carrying out several essential functions in organisms.
Also, other than proteins and lipids, carbohydrates are a vital component of the structure. They are only present on the outer side of the cell membrane, attached by covalent bonds to some lipids and proteins. Moreover, one must know that the primary physical force that organizes the lipid bilayer is “hydrophobic force.”[5]
Membrane Lipids
All membrane lipids are amphipathic – have both hydrophilic and hydrophobic ends. They constitute around 50% of the mass of most cell membranes. The proportion of lipids differs depending on the type of cell membrane. For example, the plasma membrane is composed of approximately 50% lipid and 50% proteins. Whereas the inner membrane of the mitochondria is 25% lipid and 75% protein.[5]
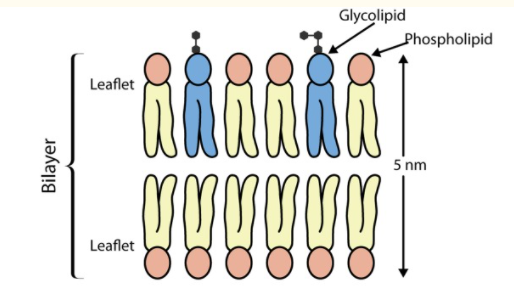
Image: An illustration of lipid bilayer showing the hydrophobic head (pink) facing an aqueous environment and the hydrophilic tail facing inwards, away from the water.
Source: Watson H. (2015). Biological membranes.[6]
Similarly, the composition of lipids in the biological membrane also differs depending on the organisms. For example, the plasma membrane of E. coli consists predominantly of phosphatidylethanolamine (constituting 80% of total membrane lipid).
In contrast, the mammalian plasma membrane comprises four different phospholipids (making up 50-60% of total membrane lipids), glycolipids, and cholesterol (which both make up the other 40%).
Below is a brief on all the three classes of lipids: Phospholipids, glycolipids, and sterols.[6]
1. Phospholipids
It consists of four components: alcohol (glycerol and sphingosine), fatty acids, phosphate, and an alcohol attached phosphate.[6]
The simplest phospholipid is phosphatidic acid, in which two fatty acid residues are esterified to the OH groups at carbon atoms 1 and 2 of glycerol-3-phosphate.
Two main types of phospholipids include glycerophospholipids and sphingophospholipids.
- Glycerophospholipid: These are phospholipids containing glycerols. Phosphatidylcholine is the most commonly found glycerophospholipid in the membrane. It has a choline molecule attached to the phosphate ring. Cells make many other types of glycerophospholipids by combining different fatty acids and head groups. Other examples are phosphatidic acid, phosphatidylethanolamine, and phosphatidylserine.[6]
- Sphingophospholipid: These molecules contain sphingosine rather than glycerols. Sphingosine is a long acyl chain with an amino group (NH2), two hydroxyl groups (OH), and a phosphocholine group (attached to the terminal hydroxyl group) at one end, forming the hydrophilic head of the sphingophospholipid’s structure. The hydrophobic fatty acid tail (attached to the amino group) forms the other end of the structure.[6]
- Phosphatidylcholine, phosphatidylserine, phosphatidylethanolamine, and sphingomyelin are the four major phospholipids, constituting more than half the mass of lipids in most mammalian cell membranes.[6]
2. Glycolipids
- The structure of glycolipids contains either glycerol or sphingosine and has sugar in place of phosphate head — different from that seen in sphingolipids.[6]
- In eukaryotic plasma membranes, it only makes up 5% of the lipid molecules in the outer monolayer.[7]
- The most complex glycolipid is ganglioside which contains oligosaccharides with one or more sialic acid moieties.[7]
3. Sterols
- Cholesterol is the principal sterol present in plasma membranes. They account for about 20% of the lipids in animal cell plasma membranes.[7]
- The structure of cholesterol consists of a hydroxyl group (which is the hydrophilic ‘head’ region), a four-ring steroid structure, and a short hydrocarbon side chain.[7]
- Plants contain sterols like stigmasterol and sitosterol in their cell membrane.
- In bacterial cell membranes, sterols are absent.
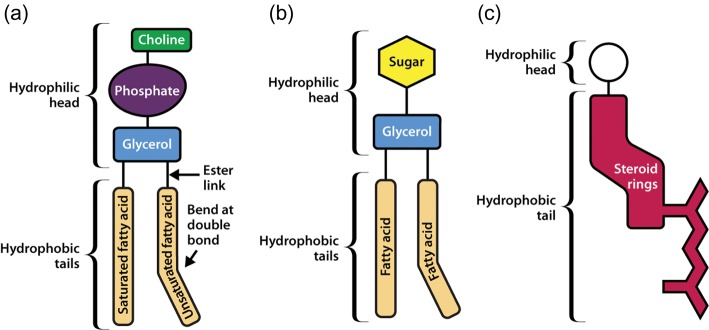
Image: An schematic representation of the structures of (a) Phosphatidylcholine, a glycerophospholipid; (b) Glycolipid; and (c) Sterol.
Source: Watson H. (2015). Biological membranes.[6]
Membrane Proteins
Membrane proteins are an integral and dynamic part of the cell membrane. They account for at least half of the mass of most membranes. Their presence is essential in the membrane for signaling, communication, and several other life processes of organisms.
There are two categories of membrane proteins based on their association with the plasma membrane:[8]
1. Intrinsic Membrane Protein
These are proteins that are completely or partially embedded within the plasma membrane. They are also known as integral membrane proteins. Most of these proteins contain hydrophobic side chains that interact with the fatty acyl group of membrane phospholipids and facilitate anchoring the proteins to the membrane.[8]
One classic example of the integral membrane protein is single pass or multipass transmembrane proteins. These proteins contain one or more membrane-spanning regions (α-helices or multiple β-strands) and regions extending in the aqueous medium on each side of the bilayer.[8]
An example of single-pass transmembrane protein is RBC glycophorins, and that of multipass transmembrane proteins is band-3 proteins or chloride-bicarbonate exchange proteins of RBCs.[8]
The integrity of the transmembrane proteins with the phospholipid bilayer makes it experimentally challenging to isolate them by simple extraction procedures.
2. Extrinsic Membrane Protein
Extrinsic membrane proteins don’t span the hydrophobic core of the membrane.[8] These proteins are either indirectly bound to the membrane through interaction with integral membrane protein or directly bound to the lipid polar head group. They are bound to membranes mainly by electrostatic and hydrogen bond interactions. These proteins are also known as peripheral membrane proteins.[8]
The extrinsic membrane proteins are localized to the cytosolic face of the membrane and play an essential role in signal transduction. These proteins include cytoskeletal proteins spectrin and actin of RBCs, and protein kinase C enzyme.[8]
The extrinsic membrane proteins can be easily isolated either by using solutions of very high or low ionic strengths, solutions of extreme pH or by gentle extraction.[8]
Membrane Carbohydrates
Carbohydrates are the third major component of plasma membranes. They are attached to the outer side of the membrane while being linked to proteins (forming glycoprotein) or lipids (forming glycolipid).[9]
They also occur as polysaccharide chains of an integral membrane, called proteoglycans. These carbohydrates are composed of 2-60 monosaccharide units (that can be either straight or branched) and coat the surface of all eukaryotic cells.[9]
- Glycoproteins: It’s composed of protein attached with one or more oligosaccharides. They participate in a wide range of cellular phenomena, including cell recognition and cell surface antigenicity.[9]
- Glycolipids: It’s a carbohydrate covalently linked to membrane lipids. They are involved in maintaining cell stability and facilitating cellular recognition.[9]
- Proteoglycans: They consist of long polysaccharide chains linked covalently to a protein core and are found outside the cell as part of the extracellular matrix. In simple words, they are proteins that are heavily glycosylated.[9] Their major functions are derived from the physicochemical characteristics of the glycosaminoglycan component of the molecule, which provides hydration and swelling pressure to the tissue enabling it to withstand compressional forces.[9]
Properties of the Lipid Bilayer
The lipid bilayer structure can be observed by using electron microscopy, x-ray diffraction, and freeze-fracture electron microscopy techniques. The techniques helped reveal the details of the membrane’s organization and its properties attributed to the lipid molecules.
Three such properties of the cell membrane include asymmetry, fluidity, formation of lipid rafts.
1. Asymmetry in the plasma membrane
The asymmetry of the plasma membrane denotes the difference in the composition of lipids in the two monolayers of a lipid bilayer. For example, in red blood cells, almost all lipid molecules with choline—(CH3)3N+CH2CH2OH—in their head group (phosphatidylcholine and sphingomyelin) are in the outer monolayer.
While almost all phospholipid molecules containing a terminal primary amino group (phosphatidylethanolamine and phosphatidylserine) are in the inner monolayer.[10]
The asymmetric lipid distribution between the two lipid monolayers also causes a charge difference between them. At neutral pH, phosphatidylserine is negatively charged, making the cytosol side of the layer more negative than the outer side.[10] The other lipids, phosphatidylethanolamine, phosphatidylcholine, and sphingomyelin, are neutral, thus, carry no charges.
2. The fluidity of the plasma membrane
The term “fluid” in the fluid mosaic model denotes the motion of the lipids in the membrane or its fluid behavior. The membrane lipid molecules possess both rotational and lateral motion. The movement depends on three factors: temperature, lipid composition, and cholesterol.[10]
- Temperature: At higher temperatures, membranes are more fluid, and at a lower temperature, they act as a viscous substance.
- Cholesterol: In eukaryotes, higher cholesterol concentration decreases the fluidity of the membrane. But, this property of cholesterol also depends on the temperature. For example, at higher temperatures, the membrane is less fluid than at lower temperatures.[10]
- Lipid Composition: Shorter chain length and higher unsaturation levels in fatty acids in the membrane increase the membrane’s fluidity.
Here’s more on the factors affecting membrane permeability and fluidity.
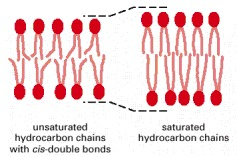
Image: An illustration of the effect of unsaturated fatty acids on membrane fluidity. The saturated fatty acids tightly pack the lipids in the membrane, thus, decreasing their fluidity.[10]
Source: Molecular Biology of the Cell. 4th edition.
3. Formation of lipid rafts in the plasma membrane
The lipid molecules in the membrane are randomly organized and connected to the neighboring fatty acids through Van der Waals attractive forces.[10] For some lipid molecules like sphingophospholipids having long and saturated fatty hydrocarbon chains, the attractive force is so strong that it links the adjacent molecules transiently in small microdomains. These microdomains are called lipid rafts.[10]
Lipid rafts are approx 70 nm in diameter and rich in sphingolipids and cholesterol. They are of two types: non-caveolar and caveolae (contain caveolin proteins). These lipids rafts are involved in signal transduction, endocytosis, and cholesterol trafficking in cells.[10]
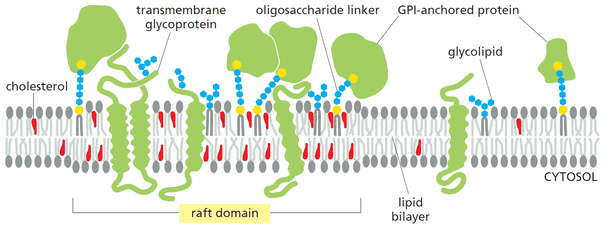
Figure: A diagrammatic representation of the lipid rafts.[7]
Source: Alberts et al. (2015) Pg. 537
Functions of Biological Membranes
- The asymmetry in the plasma membrane helps to distinguish between live and dead cells. Phosphatidylserine, usually confined to the cytosolic side of the plasma membrane lipid bilayer, rapidly translocates to the extracellular monolayer when animal cells undergo programmed cell death or apoptosis. The presence of phosphatidylserine on the cell surface serves as a signal to induce neighboring cells, such as macrophages to phagocytose the dead cell and digest it.[10]
- The glycocalyx coat formation on the cell surface protects it from damage or harm and mediates cell-cell adhesion events.[10]
- The peripheral proteins that form an extracellular matrix on the cell surface have an essential role in cell recognition.
- The plasma membrane protects the cell from the outer environment and maintains the favorable environment inside the cell.
- The biological membrane has an essential role in the transport of molecules performed by the membrane proteins. And based on these functions, membrane proteins are also classified into channel protein and carrier protein.
- Channel Protein: These proteins transport the molecules down the gradient – that is, passive transport. The channels can be gated or non-gated based on the signal required to initiate the functional response.
- Carrier Protein: These proteins can be uniport, symport, or antiport. Uniport is the transfer of molecules on only one side of the membrane, symport is the simultaneous transfer of two molecules in the same direction, and antiport is the simultaneous transfer of two molecules in opposite directions. The symport and antiport transportations are together known as co-transport. The transport of molecules through carrier proteins can be done via active transport or passive transport processes.
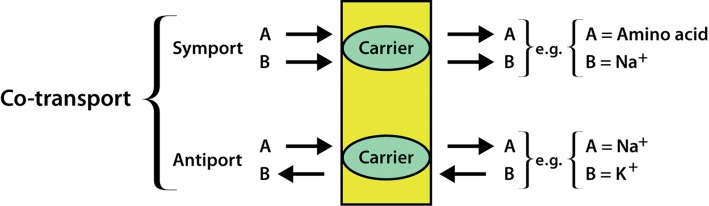
Image: An illustration of types of co-transport and their examples.[6]
Source: Watson H. (2015). Biological membranes.
Conclusion
Biological membranes are the boundary of cells that protect them from the external environment and perform essential functions required for living organisms. The properties of the cell membranes are similarly/equally shared in both prokaryotic and eukaryotic organisms.
The bilayer of the membrane is composed of three major biomolecules: lipids, proteins, and carbohydrates. Further, the advancement in biophysical techniques and availability of substantial computational power is expanding our understanding of the lipid bilayers and providing critical insights into their structures and functions.[6]
Scientists are currently working towards understanding the properties, functions, and mechanisms of the membrane proteins, which they think might be the key to fighting deadly diseases.[6]
References:
- Biological Membrane. Retrieved from https://en.wikipedia.org/wiki/Biological_membrane.
- History of Cell Membrane Theory. Retrieved from https://en.wikipedia.org/wiki/History_of_cell_membrane_theory.
- Membrane Models. Retrieved from https://en.wikipedia.org/wiki/Membrane_models.
- Cell Membrane. Retrieved from https://biology4isc.weebly.com/cell-membranes.html.
- Cooper, G. M., & Hausman, R. E. (2007). The cell: A molecular approach. 4th ed. Washington, D.C.: Sunderland, Mass.: ASM Press.
- Watson H. (2015). Biological membranes. Essays in biochemistry, 59, 43–69. https://doi.org/10.1042/bse0590043.
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2015). Molecular biology of the cell, 6th edition. New York: Garland Science.
- Lodish H, Berk A, Zipursky SL, et al. Molecular Cell Biology. 4th edition. New York: W. H. Freeman; 2000. Section 3.4, Membrane Proteins. Available from: https://www.ncbi.nlm.nih.gov/books/NBK21570/.
- Aryal Sagar (2021). Membrane Carbohydrates. Retrieved from https://microbenotes.com/membrane-carbohydrate/.
- Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002. The Lipid Bilayer. Available from: https://www.ncbi.nlm.nih.gov/books/NBK26871/