Enzymes are a group of proteins that catalyze non-spontaneous chemical reactions in any biological system. In an organism, enzymes function as a group of interconnected chemical reactions in a metabolic pathway, fulfilling a specific cellular task.
Metabolic pathways are active under normal circumstances and alter their activities in response to internal and external stimuli. Delayed response or failure to answer to changing situations can pose damaging effects to the functioning and survival of the organism.
The metabolic network is intricate and constructed to ensure that metabolic responses are specific in timing and circumstances. Regulatory enzymes contribute to the timing aspect by controlling the overall rate of a metabolic pathway. The availability of metabolites and catalytic activity of the enzymes within the pathway dictate how the cell responds to a particular event under a given circumstance.
In doing so, enzymatic reactions transpire only in suitable cellular environments and proceed at a rate appropriate to the availability of the necessary substrate or cofactors. Changes in the surrounding conditions are reflected in certain factors, promoting or suppressing the enzyme’s activity and the rate of enzymatic reactions.
These factors are:
1. Enzyme Concentration
The transient bonds between enzymes and their substrates catalyze the reactions by decreasing the activation energy and stabilizing the transition state. Given the exceeding amount of substrates and the necessary cofactors, enzymatic reactions possessing higher enzyme concentrations will reach equilibrium before those with the same enzyme but at lower concentrations.
Simply put, higher enzyme concentration indicates that more enzyme molecules are available to process the substrate. The high levels of enzyme-substrate complex result in a higher initial catalytic rate, which gives the reaction a headstart in the shift toward reactant-product equilibrium.
2. Substrate Concentration
The enzyme catalytic activity occurs when a geometrically and electronically complementary substrate can access the enzyme’s catalytic or active site. There, the active residues transiently bond with the substrate, catalyzing the transformation of the substrate into a product. Thus, the more substrate-occupied active sites, the higher the catalytic activity and the faster the shift toward enzyme-product equilibrium.
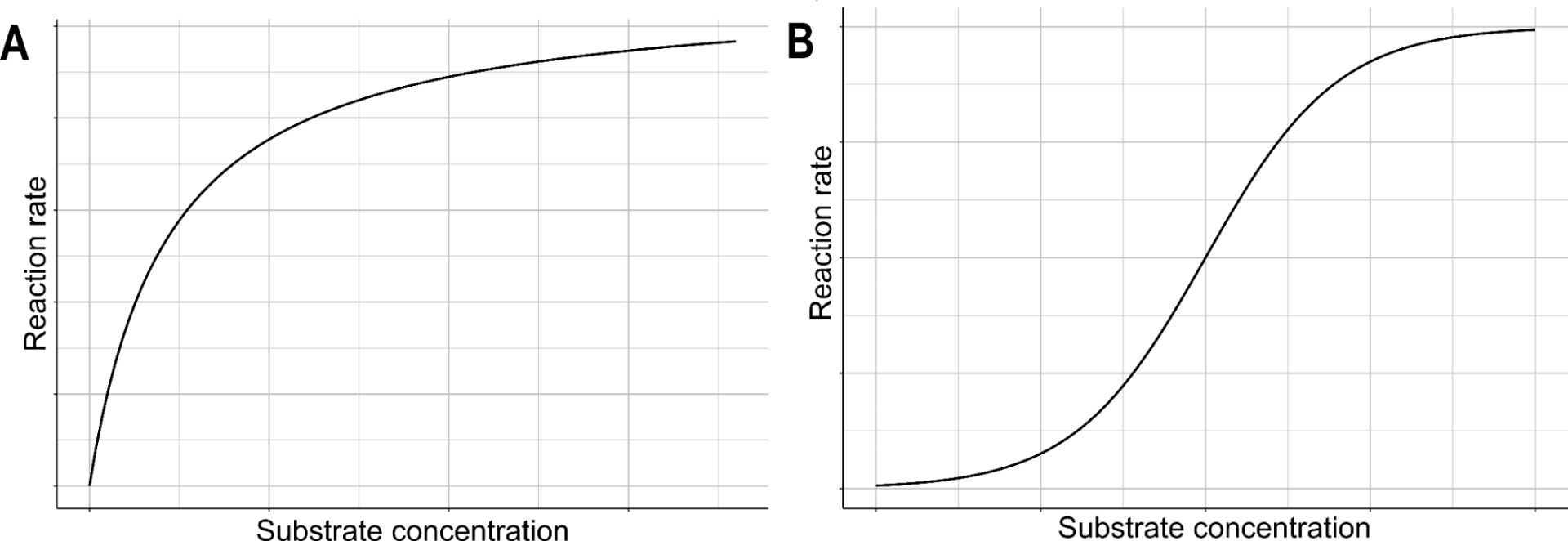
Most enzymes follow the Michaelis-Menten kinetics, which describes the relationship between enzyme activity and substrate concentration in two stages. At the initial stage, the relationship between the two is a linear association and plateaus when the number of unbound active sites decreases.
Another group of enzymes, allosteric enzymes, display a sigmoidal kinetic. Initially, the relationship between the rate of an allosteric enzyme-catalyzed reaction is exponential. However, this becomes linear as the catalysis progresses and finally plateaus when the number of substrate-bound enzymes becomes saturated.
Figure 1: The relationship between substrate concentration and the rate of enzyme-catalyzed reaction follows the Michaelis-Menten kinetic in most enzymes (A) but a sigmoid curve in allosteric enzymes (B).
3. pH Value
As a chain of amino acids, proteins such as enzymes contain electrical charges from the sequence of their amino acid residues. Most amino acids in the chain are the basis for the intramolecular interactions that give the enzyme its three-dimensional structure. Few others act as functional residues at the enzyme’s active site.
Altogether, the amino acids determine the substrate specificity and restrict the enzyme activity only to a narrow range of pH. Most enzymes function optimally in slightly acidic or basic pH. However, a few enzymes are native to extreme acidic or basic environments; hence, most active in these pH ranges.
For this reason, a change in the pH value, either acidic or basic, affects the ionization of amino acid residues, leading to changes in the three-dimensional structure of the enzyme. The alteration in the enzyme conformation affects its interaction with its substrate, thus reducing its activity.
Another effect of pH change is in the enzyme’s catalytic capability. In acid-base and covalent catalysis mechanisms, pH change can hinder or suppress catalytic activity. In extreme cases, it can denature the enzyme, destroy its three-dimensional structure, and render it permanently non-functional.
Content
1. ɑ-Amylase
In saliva, amylase breaks down most polysaccharides in human diets.
6.4 – 7.0
6.6
Content
2. Pepsin
Pepsin is one of the many proteases found in the stomach’s gastric juice. It hydrolyzes peptide bonds in the protein’s amino acid chains.
1.5 – 4.5
2
Content
3. Trypsin
Found in the small intestine, trypsin is another protease that digests proteins.
7.5 – 8.5
7.8
Content
4. Alkaline Phosphatase (ALP)
ALP catalyzes the removal of phosphate groups from its substrate. It is found in all human tissue and is most abundant in the intestine and placenta.
8 – 10
10
Table 1: Examples of enzymes in humans, their function, pH range, and optimal pH
4. Temperature
In the same way that pH affects enzymes, temperature also influences the stability of their intramolecular bonds. For this reason, enzyme activity is generally more active at their optimal temperature.
Nonetheless, a few degree shifts from the optimal temperature only cause a minor decrease in the enzyme activity.
Content
1. Thermococcus hydrothermalis
Prokaryotic archaea found in the East Pacific hydrothermal vent
5.5
85°C
Content
2. Sulfolobus solfataricus
Prokaryotic archaea found in sulfur-rich volcanic fields
3.0
80°C
Content
3. Halomonas meridiana
Gram-negative bacteria found in Antarctica salt lake
7.0
37°C
Content
4. Pseudoalteromonas haloplanktis
Fast-growing bacteria found in Antarctic seawater
7.6
4°C
Table 2: Examples of optimal pH and temperature of ɑ-Amylase from selected organisms.
A slight increase in the temperature can speed up the reaction rate as the reactants acquire more kinetic energy. Significant deviations from the optimal temperature, however, significantly reduce the enzyme activity. Extreme high temperatures can destroy the intramolecular bonds and the enzyme conformation, rendering it permanently non-functional.
Low temperature decreases the kinetic energy of the system and reduces the reaction rates. Enzyme activity declines as the temperature gradually fall below the optimal point. Unlike the case of high temperature, low temperature does not necessarily result in permanent enzyme denaturation, and the enzyme activity may be restored once the temperature rises to the optimal range.
Since enzymes generally exist in aqueous solutions, a decrease in temperature upsets the nature of its interaction with water, reducing its solubility and causing the enzyme to unfold – this ultimately inactivates the enzyme.
However, when the temperature falls below the melting point of water (0°C or 32°F), it leads to the formation of ice crystals that can irreversibly damage the proteins. The same effect is also seen when frozen enzymes are thawed. The freeze-thaw damage can be avoided by minimizing freeze-thaw cycles, freezing or thawing duration, and adding additives like sucrose or glycerol to the protein solution.
5. Effector or Inhibitor
Many enzymes require non-substrate and non-enzyme molecules to regulate or initiate their catalytic function. For example, certain enzymes rely on metal ions or cofactors to establish their catalytic activity. Many rely on effectors to activate their catalytic activities, promote or inhibit their successive binding to the substrates, as seen in allosteric enzymes.
Along the same line, inhibitors may bind to the enzyme or its substrate to inhibit the ongoing enzymatic activity and prevent successive catalytic events. The effect on enzyme activity is irreversible when the inhibitors form strong bonds to the enzyme’s functional group, leaving the enzyme permanently inactive.
In contrast to irreversible inhibitors, reversible inhibitors only render the enzymes inactive when bound to the enzyme. Competitive inhibitors compete with the substrates for binding to the residues of the enzyme functional group at the catalytic sites. Other types of inhibitors do not bind to the catalytic site, but they bind to the non-substrate binding allosteric site.
If an inhibitor binds to the enzyme concurrently with the enzyme-substrate binding, it is non-competitive. If an inhibitor binds only to a substrate-occupied enzyme, it is uncompetitive.
In Conclusion
All in all, enzymes play a vital role in metabolic responses, shaping how cells and organisms mature and adapt. Enzyme and substrate concentrations influence the reaction rate. Factors such as pH, temperature, effectors, and inhibitors modify the enzyme conformation, altering its catalytic activity.
Altogether, they reflect the current metabolic situations and trigger changes in the inherent characteristics of the enzyme and its interaction to promote or impede enzymatic reactions.
References:
Voet D, Voet JG and Pratt CW, Fundamentals of Biochemistry, 2nd edition. New Jersey: John Wiley & Sons; 2006.
Boyer R, Concepts in Biochemistry, 3rd edition. New Jersey: John Wiley & Sons; 2006.
Punekar, N S. 2018. “Hallmarks of an Enzyme Catalyst.” In ENZYMES: Catalysis, Kinetics and Mechanisms, 43–51. Singapore: Springer Singapore. doi:10.1007/978-981-13-0785-0_5.
Marini I. Discovering an accessible enzyme: Salivary α-amylase : Prima digestio fit in ore: A didactic approach for high school students. Biochem Mol Biol Educ. 2006;33(2):112-116. doi:10.1002/bmb.2005.494033022439
Piper DW, Fenton BH. pH stability and activity curves of pepsin with special reference to their clinical importance. Gut. 1965;6(5):506-508. doi:10.1136/gut.6.5.506
Lam MPY, Lau E, Liu X, Li J, Chu IK. Sample Preparation for Glycoproteins. In: Comprehensive Sampling and Sample Preparation. Elsevier; 2012:307-322. doi:10.1016/B978-0-12-381373-2.00085-5
Linden G, Alais C. Alkaline phosphatase in human, cow and sheep milks: molecular and catalytic properties and metal ion action. Ann Biol Anim Biochim Biophys. 1978;18(3):749-758. doi:10.1051/rnd:19780412
Vieille C, Zeikus GJ. Hyperthermophilic Enzymes: Sources, Uses, and Molecular Mechanisms for Thermostability. Microbiol Mol Biol Rev. 2001;65(1):1-43. doi:10.1128/MMBR.65.1.1-43.2001
Mehta D, Satyanarayana T. Bacterial and Archaeal α-Amylases: Diversity and Amelioration of the Desirable Characteristics for Industrial Applications. Front Microbiol. 2016;7. doi:10.3389/fmicb.2016.01129
Feller G, Payan F, Theys F, Qian M, Haser R, Gerday C. Stability and structural analysis of alpha-amylase from the antarctic psychrophile Alteromonas haloplanctis A23. Eur J Biochem. 1994;222(2):441-447. doi:10.1111/j.1432-1033.1994.tb18883.x
James SR, Dobson SJ, Franzmann PD, McMeekin TA. Halomonas meridiana, a New Species of Extremely Halotolerant Bacteria Isolated from Antarctic Saline Lakes. Syst Appl Microbiol. 1990;13(3):270-278. doi:10.1016/S0723-2020(11)80198-0
Georlette D, Blaise V, Collins T, et al. Some like it cold: biocatalysis at low temperatures. FEMS Microbiol Rev. 2004;28(1):25-42. doi:10.1016/j.femsre.2003.07.003.
Cao E, Chen Y, Cui Z, Foster PR. Effect of freezing and thawing rates on denaturation of proteins in aqueous solutions. Biotechnol Bioeng. 2003;82(6):684-690. doi:org/10.1002/bit.10612.