




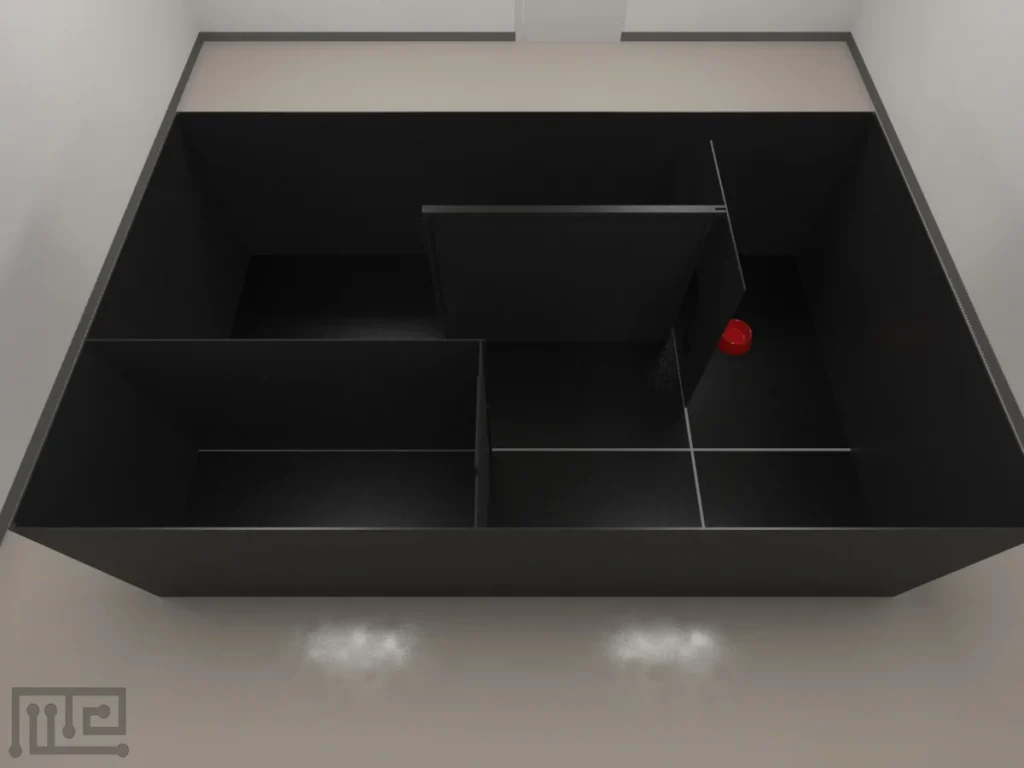
MazeEngineers offers custom-built behavioral mazes at no extra cost—designed to fit your exact research needs. Eliminate reproducibility issues from poor sizing or lingering scent cues with precision-engineered, modular, and smart mazes that adapt in real time to animal behavior. Publish new protocols, run adaptive experiments, and push the boundaries of behavioral science.
























































































































Reviews
There are no reviews yet.